酵母氨酸还原酶影响真菌中苦马豆素的合成
2019-06-17赵利娜
赵利娜,卢 萍
(内蒙古师范大学 生命科学与技术学院,内蒙古 呼和浩特 010022)
1 引言
小花棘豆(OxytropisglabraO.glabra)是生长于荒漠草原低湿地隶属于豆科(Leguminose)棘豆属(OxytropisDC.)的多年生草本植物,与黄芪属(AstragalusstragalusA.stragalus)统称为疯草,整个生长周期均含有毒生物碱-苦马豆素(swainsonine,SW)[1]。SW结构类似甘露糖的阳离子半椅式构象,与甘露糖苷竞争性结合,对α-甘露糖苷酶的活性具有抑制作用,使牲畜细胞功能出现紊乱[2],发生中毒症状,严重者致死,给草原上畜牧业带来极大的损失[3]。研究者对含有SW的植物体进行研究,发现其毒性是由于植物体分离培养的内生真菌合成的[4]。酵母氨酸还原酶基因(saccharopine reductase gene,sac)编码酵母氨酸还原酶(saccharopine reductase,Sac)[5],是合成SW代谢途径中重要前体化合物[6,7]。本文主要对豆类丝核菌(Rhizoctonialegumimicola,R.legumimicola)、金龟子绿僵菌(Metarhiziumanisopliae,M.anisopliae)、稻瘟病菌(Magnaporthegrisea,M.grisea)以及疯草内生真菌(Alternariaoxytropis,A.oxytropis,)等微生物合成苦马豆素途径中sac对其代谢影响进行综述。
2 酵母氨酸还原酶
酵母氨酸还原酶基因编码酵母氨酸还原酶,在微生物中赖氨酸由二氨基庚二酸和α-氨基己二酸两条途径分别合成,在α-氨基己二酸途径中,α-氨基己二酸半醛 Sac 酵母氨酸→ 赖氨酸,酵母氨酸是合成赖氨酸的中间产物[5]。反应过程中Sac的热稳定性较差,在46 ℃时,酶失活发生双向反应,缓冲液pH值为中性时,发生正向反应,生成酵母氨酸;缓冲液pH值为碱性时,发生逆向反应,生成谷氨酸[8]。
1988年Pearson等在酿酒酵母Saccharomyces cerevisiae(S.cerevisiae)中克隆到sac[9]。2000年Johansson等在M.grisea中克隆到sac(GenBank登陆号:AF144424),与S.cerevisiae的sac的序列比对一致度达63%,并在M.grisea中提取到Sac粗酶对其进行了纯化[10]。随后2006年接着对Sac利用分子置换法发现其单体酶分别由结构域I的折叠变体Rossmann(用于结合NADPH)、结构域Ⅱ折叠成α/β结构(用于结合糖蛋白并参与二聚体形成)和结构域Ⅲ为全螺旋折叠(用于与底物结合时,关闭酶的活性位点)三部分组成[12]。对S.cerevisiae和M.grisea的Sac氨基酸结构进行比较,发现结构域II具有最高数量的保守残基,在底物结合和空间定向中结构域Ⅰ和Ⅲ中起到重要作用[11]。2008年Albang等在产黄青霉菌Penicillium chrysogenum中克隆到sac序列(GenBank登陆号:XM002564566),随后利用同源重组获得了sac失活突变株[13,14]。
2010年Mukherjee等利用简并PCR在A.oxytropis中克隆到sac的cDNA序列(GenBank登陆号:HQ010362)[15]。卢萍课题组在2014年对内蒙古的O.glabra体外分离得到内生真菌,利用RACE获得sac的cDNA序列(NCBI登录号:KJ944635),简并PCR克隆到sac的DNA序列(NCBI登录号:KY052048),与从美国A.oxytropis克隆得到的sac的cDNA序列一致度为82.3%[16]。随后构建sac缺失载体,获得sac基因缺失突变株M1[17],通过Southern杂交,检测到M1中的sac中间片段已被敲除,且hph整合到M1的基因组中[18]。成功构建A.oxytropis内生真菌sac互补表达载体,获得A.oxytropis内生真菌sac互补株C1[19]。
3 苦马豆素的性质及其来源
SW又名吲哚兹定三醇(indolizidine triol),是一种多羟基吲哚兹定的有毒生物碱,化学式为C8H15NO3[20]。因化学结构与甘露糖相识度高,强力抑制α-甘露糖苷酶的活性,致使动物细胞溶酶体内的α-甘露糖苷大量贮积,细胞合成糖蛋白发生障碍,细胞出现空泡变性[21]。牲畜大量采食影响神经系统的发育、肝功能损伤,免疫系统絮乱,严重者致死。近年由于SW可促进骨髓细胞和免疫细胞的增生以及细胞分化,也可以直接对肿瘤细胞进行抑制,改变细胞表面的糖基化,配合化疗药物促进细胞杀伤肿瘤细胞,使肿瘤细胞凋亡,广泛用于机体增强免疫功能和癌症的治疗,目前SW主要从植物体内提取、化学人工合成和产SW的真菌中提取[22]。
Colegate等首次于1979年在灰苦马豆(Swainsonacanescens)植物体中分离提取到SW,并将其命名为SW[23]。随后Molyneux等在绢毛棘豆和黄芪中检测到高浓度SW,认为致使牲畜中毒主要是因为其植物体含SW[24]。1989年曹光荣等从黄花棘豆(OxytropisochrocephalaBunge)中分离到SW,并证实SW致使细胞形成空泡化,使其失去功能后逐渐死亡[25]。1995年Sim等从M.anisopliae分离到SW,利用酶分析法测得SW的水平[26]。2003年Braun等在绢毛棘豆Oxytropissericea、斑荚膜黄芪(Astragalusmollisimus)和蓝伯氏棘豆(OxytropislambertiiO.lambertii)3种植物体的不同部位,通过体外分离培养得到内生真菌,经过形态学和分子生物学鉴定为Embellisia,并认为体外培养该内生真菌可合成SW[27]。卢萍等2004年在内蒙古O.glabra植物体中体外分离培养到内生真菌[28],观察形态以及5.8S rDNA/ITS序列比对,一致度达99.8%,鉴定为Embellisia,并发现小花棘豆毒性来源体外培养的内生真菌Embellisia,含该内生真菌的小花棘豆宿主均含SW,不含该内生真菌的宿主测不出SW[28]。Ralphs等在2008年从美国疯草不同属植株体中均分离到含SW的内生真菌Embellisia[30]。Cook在2009年对不同种的群绢毛棘豆中的SW水平进行研究发现,SW水平高的种群植株分离到的内生真菌多,而SW水平低的种群分离到的内生真菌少[31]。
2009年Pryor等对从O.lambertii和O.kansuensis中分离到的内生真菌进行研究,认为其分生孢子有较厚的横壁,形状为倒棒状或卵形,但由于分生孢子形状为波浪形,有分支芽管,又经5.8S rDNA/ITS1-ITS2序列测序分析比对,将内生真菌Embellisia修订为Undifilum[32],2014年Woudenberg等又将Undifilum归到Alternaria[33-34],小花棘豆内生真菌的学名修订为Alternaria oxytropis(A.oxytropis)。利用qPCR法对O.glabra植物体的不同部位中内生真菌数量及SW水平检测分析,发现不同植物体部位内生真菌数量不同,SW水平也不同[36]。
4 Sac影响真菌中SW的合成
1988年Harris等研究R.legumimicola中合成SW的生物代谢途径,提出SW的代谢途径可能为:赖氨酸→酵母氨酸→L-2-氨基己二酸半醛→1,6-二六氢哌啶酸(P6C)→哌啶酸→1-酮基吲哚里西啶,随后1-酮基吲哚里西啶通过两条途径从顺式羟基吲哚里西啶和反式羟基吲哚里西啶分别的生成根霉菌氨(S型-C)和SW(R型-C)[37]。1990年Wickwire等对合成SW途径中的哌啶酸进行研究,发现赖氨酸→酵母氨酸→P6C→哌啶酸,哌啶酸是由赖氨酸生成的,且哌啶酸是合成SW的前体物[38](图1)。
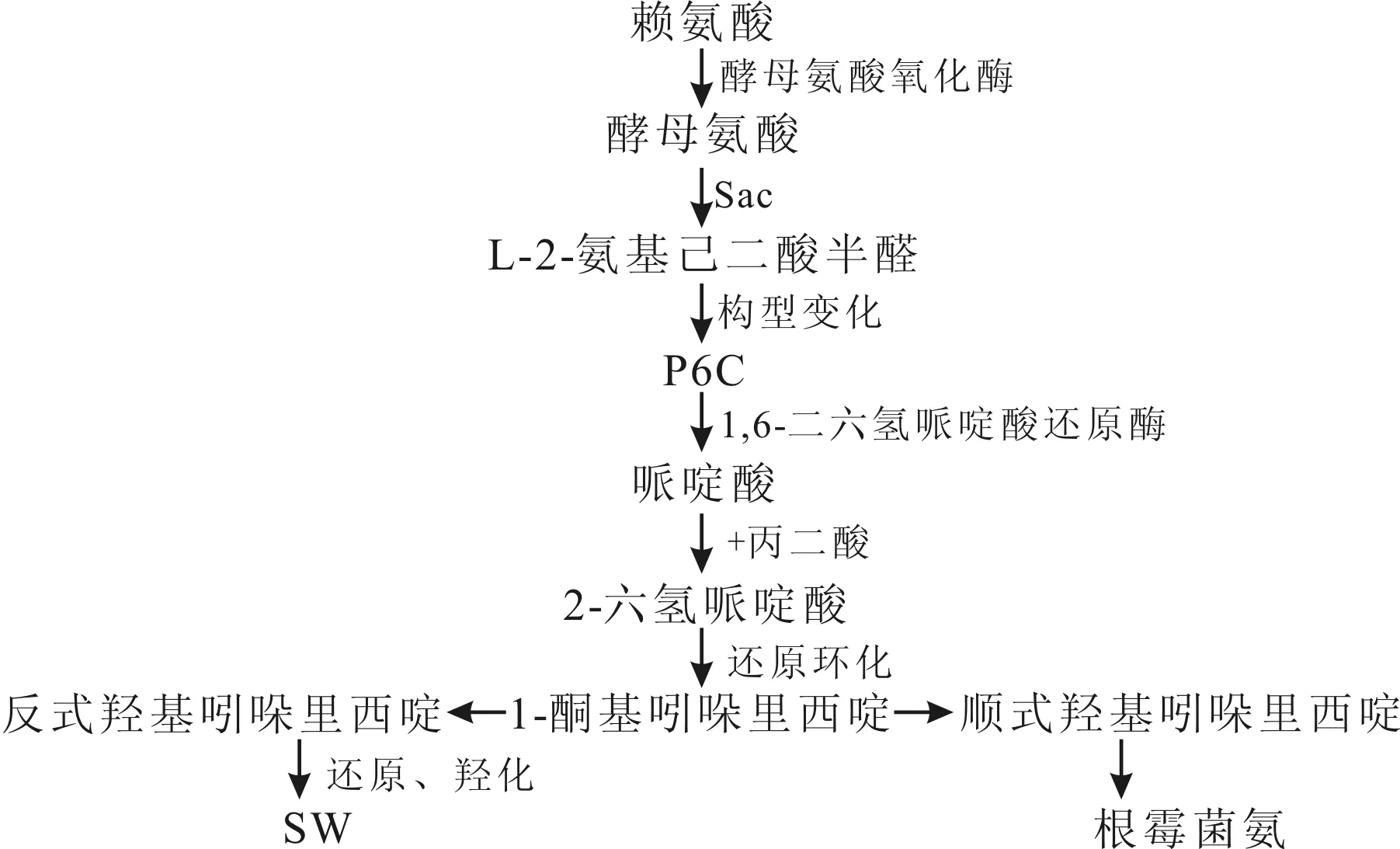
图1 豆科丝核菌中SW合成代谢途径
1997年Sim等在M.anisopliae中分离得到SW,推测SW代谢途径如图2, SW分别从不同两条路线生成P6C后,再合成SW,而赖氨酸是SW的前体化合物,在培养基中添加赖氨酸,菌丝体中哌啶酸和赖氨酸积累,添加2-氨基乙基半胱氨酸(AEC),菌体中SW水平降低,但酵母氨酸量增多,菌体需要大量赖氨酸与AEC竞争,使酵母氨酸合成赖氨酸,生成SW[39]。
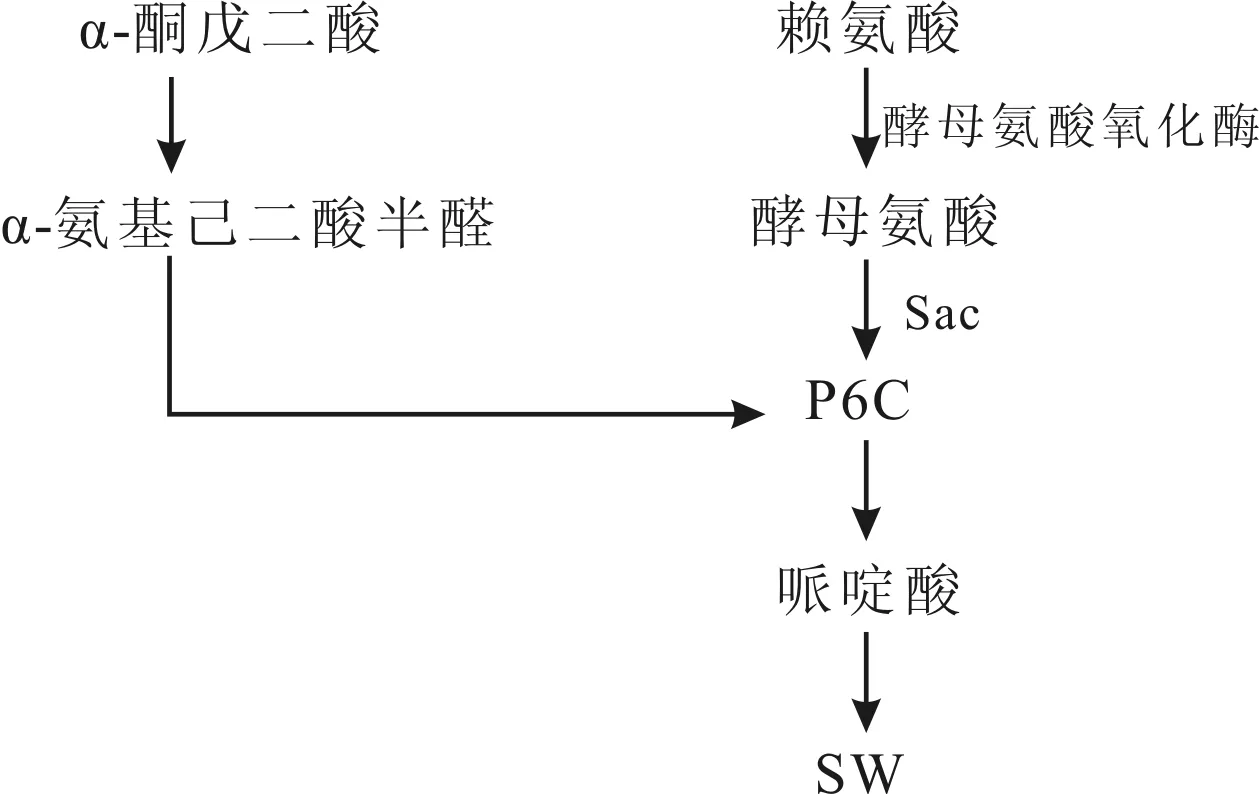
图2 金龟子绿僵菌中SW合成代谢途径
Mukherjee等的研究表明,敲除 A.oxytropis 内生真菌中的sac,SW随着哌啶酸的增加而增加,Sac的活性影响SW合成[40]。2012年杨国栋等对赖氨酸上的C、N原子标记进行示踪实验,发现C、N进入到了SW,在培养基内分别添加α-酮戊二酸、赖氨酸以及哌啶酸,发现在培养基内添加了化合物的内生真菌中SW水平高于未添加的实验对照组,且添加哌啶酸对合成SW的影响最大[40]。随后发现添加化合物不仅影响合成SW水平也影响内生真菌的生长速率[42]。2016年卢萍等利用HPLC-MS法检测到M1中的酵母氨酸的含量低于OW7.8,赖氨酸的含量没有明显变化[43],接着课题组在OW7.8和M1培养基内添加不同浓度a-氨基己二酸、赖氨酸、哌啶酸、培养不同天数的内生真菌中的SW水平进行检测,发现添加前体物均促进OW7.8和M1内生真菌中SW的合成,添加化合物的OW7.8的SW含量高于添加化合物的M1[7]。
2017年路浩等在基因组测序了 A.oxytropis 内生真菌,对内生真菌中的基因组进行了草图组装,共发现含蛋白质编码基因有11057个,其中发现164个基因可对酶进行编码,推测在合成SW途径中酵母氨酸还原酶、酵母氨酸氧化酶、吡咯啉-5-羧酸还原酶参与合成SW[44]。2017年Ren等研究 A.oxytropis 的SW合成途径,推测SW可能是由P6C和P2C两条途径合成,其中,P6C合成途径是由a-氨基己二酸 酵母氨酸脱氢酶 P6C→哌啶酸→1-酮基-吲哚里西啶→SW[45];而P2C合成途径是由赖氨酸→α-己酮酸→P2C→哌啶酸→1-酮基-吲哚里西啶→SW[46],其合成SW途径中的细节步骤及参与的催化酶尚不清晰,还需继续深化研究[47]。A.oxytropis内生真菌中SW合成代谢途径如图3。
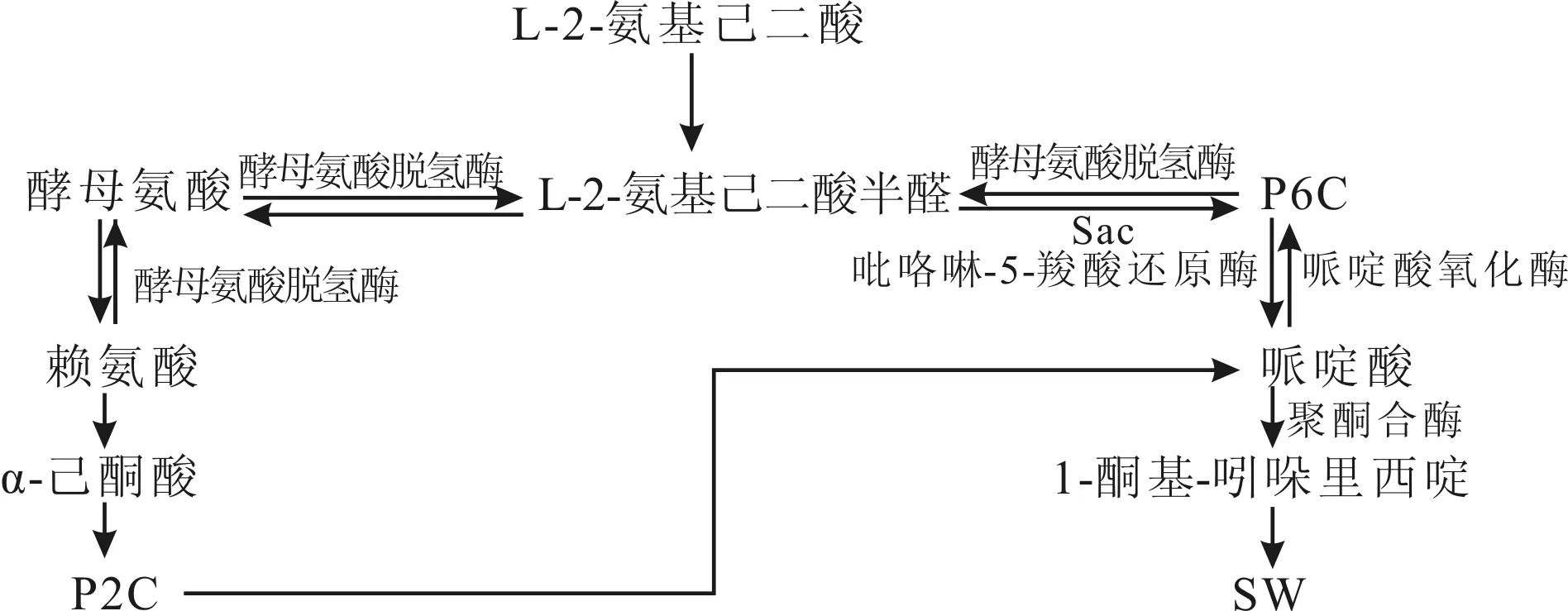
图3 内生真菌合成SW的代谢途径
5 展望
疯草内生真菌含有主要有毒成分SW,危害着草原以及畜牧业的发展,有学者发现SW具有较强的抗肿瘤作用,因而成为现下研究热点。Sac敲除突变株M1的SW水平下降,Sac促进SW的合成。未来可通过转录组分析、中间代谢物检测和关键基因的克隆与功能研究探索内生真菌SW的生物合成途径的详细细节,深入揭示SW合成的分子机制,对于草原上疯草植物小花棘豆植株进行遗传改良和生态治理,实现对其综合利用。
