日本结缕草ZjZFN1基因对拟南芥的转化及其耐旱性分析
2019-04-23姜红岩滕珂檀鹏辉尹淑霞
姜红岩,滕珂, 檀鹏辉,尹淑霞*
(1.北京林业大学草坪研究所,北京 100083;2.北京市农林科学院,北京草业与环境研究发展中心,北京 100097)
锌指蛋白是植物中一个转录因子大家族,主要由半胱氨酸(Cys)和组氨酸(His)组成,结合锌离子折叠成手指状结构的蛋白[1]。 锌指蛋白被分类为几个亚家族,例如C2H2,CCCH,C2HC,C2HC4和C5[2]。目前为止,C2H2是植物中锌指蛋白最丰富的家族之一,在拟南芥(Arabidopsisthaliana)和水稻(Oryzasativa)中分别鉴定出176和189 个C2H2锌指蛋白[3-4]。这些C2H2锌指蛋白的功能在模式植物和作物中已经得到证明,研究表明它们在植物发育、植物对生物和非生物胁迫的响应中起关键作用。例如,ZAT7或ZAT10的组成型表达增强了拟南芥对盐度的耐受性[5-6]。ZAT18[4]和GsZFP1[7]的过表达增强了拟南芥的干旱耐受性,而GmZFP3[8]负调控拟南芥的抗旱性。在盐芥(Thellungiellasalsuginea)中[9],高盐、干旱能诱导ThZF1基因表达,在盐胁迫下拟南芥突变体azf2过表达ThZF1的生长状况与野生型拟南芥无明显差异。越来越多的证据表明,与许多栽培作物品种相比,一些牧草和草坪草对各种非生物和生物胁迫的耐受性更强[10]。然而C2H2锌指蛋白在胁迫响应中的功能和潜在的转录调控机制相关的基因仍不清楚,尤其是在草坪草中。
日本结缕草(Zoysiajaponica)是耐旱性最强的草坪草种之一,普遍应用于运动场、园林绿化和水土保持等方面[11]。目前为止,对日本结缕草进行的研究主要集中在评估不同品种之间的非生物胁迫耐受性,报告生理机制以及分子标记的发展[12-13]。由于遗传资源有限,日本结缕草耐旱的分子机制尚不清楚。单个C2H2锌指蛋白的功能研究不仅可以更好地了解其在植物非生物胁迫响应中的功能,还可以深入了解植物在胁迫条件下发生的潜在信号传导过程。因此,本研究旨在获得转ZjZFN1基因拟南芥,并对其进行干旱处理,观察发芽和生长状况,并且通过实时荧光定量技术对干旱处理后转基因拟南芥中抗旱相关基因的表达模式进行研究,为进一步探究C2H2型锌指蛋白ZjZFN1在干旱胁迫中的功能及分子机制提供参考。
1 材料与方法
1.1 材料
本实验所用日本结缕草草种为‘Meyer’,由江苏省中国科学院植物研究所刘建秀研究员惠赠。实验所用的野生型拟南芥为本实验室保存,种植于人工气候箱(RXZ-380D-LED),培养条件为24/22 ℃(昼/夜),65%湿度,16 h光照。反转录试剂盒、PrimeSTAR Max DNA Polymerase、pMD19-T载体、SYBR Mix均购自TaKaRa公司;大肠杆菌感受态DH5α购自北京全式金生物技术有限公司;2×GoldStar MasterMix购自康为世纪公司;Seamless Assembly Cloning Kit购自中美泰和生物公司;GUS染色试剂盒购自奥博来公司。
1.2 方法
1.2.1ZjZFN1基因及其启动子表达载体的构建及拟南芥转化 2017年7月以健康生长3个月的日本结缕草为材料,利用‘CTAB’方法[14-15]和‘Trizol’方法[16]分别从叶片中提取基因组DNA和总RNA。以提取的DNA为模板,利用染色体步移的方法,以pMD19-T为载体,获得ZjZFN1启动子序列。将提取的RNA反转录为cDNA,并利用RACE技术进行3′/5′ RACE扩增,以pMD19-T为载体,获得ZjZFN1基因的ORF。以pMD-ZjZFN1-Pro/pMD-ZjZFN1质粒为模板,利用PrimeSTAR Max DNA Polymerase进行PCR扩增,分别用NcoⅠ/Bgl Ⅱ单酶切pCAMBIA1391Z/3302Y载体,通过Seamless Assembly Cloning Kit将纯化后PCR产物与酶切产物进行连接。1391Z-ZjZFN1 pro::GUS、3302Y-ZjZFN1载体构建成功后,利用冻融法将其转入农杆菌GV3101感受态细胞中,挑取阳性克隆菌落进行PCR鉴定(表1)。将含有表达载体的农杆菌重悬于侵染液中,使用花序侵染[17]将野生拟南芥的花序浸泡在侵染液中30 s,放于培养箱中暗培养12 h,一周后再次侵染,侵染2~3次。
1.2.2GUS基因表达检测 将收获的转ZjZFN1-Pro的拟南芥种子播种于含有20 mg·L-1潮霉素的MS培养基上进行筛选。拟南芥生长一个月左右,提取拟南芥叶片的DNA,以其为模板,1391Z-ZjZFN1 pro::GUS为阳性对照,野生型拟南芥为阴性对照,进行PCR检测。将抗性株系继续培养并筛选,直至获得T3代转基因植株,用于GUS化学组织染色。
将生长2周后的转基因拟南芥移至含有200 mmol·L-1甘露醇的MS培养基上生长,3 d后提取转基因和野生型拟南芥叶片的RNA,并反转录为cDNA,以其为模板,以qGUS-F/qGUS-R为引物进行实时荧光定量检测,以拟南芥AtUBQ10基因(NM_116771)作为内参基因。采用2-ΔΔCT法[18]计算基因的相对表达量。
1.2.3转基因拟南芥的筛选及转基因植株的检测 将收获的转ZjZFN1的拟南芥种子直接播种于含有草炭、蛭石和珍珠岩(体积比为3∶3∶1)的营养基质中,生长两周后外施60 mg·L-1草铵膦,筛选转基因拟南芥株系。利用CTAB法[14-15]提取转基因型和野生型拟南芥基因组DNA,通过基因组PCR验证阳性转基因植物,以gDNA为模板,并以3302Y-F/3302Y-R为引物进行PCR鉴定。对表现出对草铵膦100%抗性的株系进行保种并繁殖,直至获得T3代转基因植株,用于进一步的表型观察和后续实验。
1.2.4转基因拟南芥的干旱处理及抗旱性测定 将转基因拟南芥和野生型拟南芥的种子用70%乙醇和1%次氯酸钠灭菌,然后分别在含有和不含有200、300 mmol·L-1甘露醇的MS培养基上播种。12 d后,观察发芽情况及其表型。为了测定耐旱性,将3周龄幼苗移植到含有草炭、蛭石和珍珠岩(体积比为3∶3∶1)的营养基质中,转移至光照培养箱中。对其进行干旱处理21 d后观察植株表型变化。对干旱处理前和干旱处理第21天时的叶片进行取样,放于液氮中速冻,之后放于-80 ℃冰箱保存,用于脯氨酸、丙二醛(MDA)含量的测定和抗旱相关基因表达分析。
1.2.5转基因拟南芥抗旱相关基因的表达分析 利用Trizol法[16]提取上述所取样品的总RNA,经检验合格后以其为模板,通过反转录试剂盒获得cDNA。通过qRT-PCR技术,对转基因和野生拟南芥中ZjZFN1基因及干旱处理后抗旱相关的几个基因抗坏血酸过氧化物酶(ascorbate peroxidase,APX)基因、过氧化物酶(peroxidase,POD)基因、超氧化物歧化酶(superoxide dismutase,SOD)基因、吡咯啉-5-羧酸合成酶(pyrroline-5-carboxylate synthase,P5CS)基因以及晚期胚胎发生丰富蛋白(late embriogenesis abundant protein,LEA)基因进行实时荧光定量检测,以拟南芥AtUBQ10基因(NM_116771)作为内参基因。采用2-ΔΔCT法[18]计算基因的相对表达量。
1.3 统计分析
采用WPS中的Excel 2019软件作图,采用SPSS 23.0软件进行统计分析。
2 结果与分析
2.1 ZjZFN1启动子活性分析
将转ZjZFN1-Pro::GUS的拟南芥植株进行GUS组织化学染色,组织化学染色显示转ZjZFN1-Pro::GUS拟南芥植株的叶片呈现蓝色(图1A),表明ZjZFN1的启动子能够驱动GUS基因在叶片中的表达。在GUS染色结果的基础上,用200 mmol·L-1的甘露醇处理转ZjZFN1-Pro::GUS植株,处理3 d后,提取叶片的RNA并反转录成cDNA,对GUS基因进行荧光定量检测。荧光定量表达分析表明,200 mmol·L-1甘露醇处理后GUS基因的表达水平高于对照中的表达水平(图1C)。通过对干旱处理前后的拟南芥植株进行GUS染色(图1A,B),结果表明200 mmol·L-1甘露醇处理后的GUS染色强于处理前的染色结果,与荧光定量结果相符。
2.2 转基因拟南芥植株的鉴定
通过花序侵染法,转化拟南芥,获得T0代转基因拟南芥种子。通过将T0代种子播种于营养基质中并外施草铵膦,筛选抗性植株。具有抗性的拟南芥植株正常生长,保持绿色,而没有抗性的拟南芥植株逐渐枯萎死亡。提取具有草铵膦抗性的拟南芥基因组的DNA,进行PCR检测。PCR产物经电泳检测显示(图2),抗性植株DNA的PCR产物与目的条带大小一致。初步证明,目的基因转化拟南芥成功。获得T1代转基因植株后,收取T1代种子,继续筛选。最终获得T3代纯合转基因株系。因为株系ZFN-2和ZFN-17在36个T 3转基因株系中呈现最高的ZjZFN1转录水平,所以选择它们作为表型观察的代表株系。
2.3 干旱胁迫下转基因拟南芥的生长状况
将转基因株系ZFN-2、ZFN-17和WT播种于MS培养基上生长,一周之后3种株系之间的发芽率没有明显差异性(图3A)。而在添加了甘露醇的MS培养基上,转基因株系和WT的发芽率差异显著,WT的发芽率最高(图3B)。在12 d后,ZFN-2、ZFN-17和WT在MS培养基和添加了200 mmol·L-1甘露醇的MS培养上生长正常,发芽率基本一致,均长出了幼叶,未含有甘露醇的MS上的拟南芥子叶较大;而在300 mmol·L-1甘露醇的MS培养基上WT的发芽率明显高于转基因拟南芥的,仅有WT长出了绿色的子叶(图3C)。将正常生长3周后的拟南芥移栽到基质中生长,干旱处理21 d后,ZFN-2、ZFN-17植株明显比WT的矮小,叶片失去色泽,萎蔫面积多于WT(图3D)。

图1 转基因拟南芥中ZjZFN1启动子活性分析Fig.1 Activity analysis of ZjZFN1 promoter in transgenic Arabidopsis A,B:转基因拟南芥中的GUS染色 GUS staining in transgenic Arabidopsis.A: 200 mmol·L-1甘露醇处理前 Before treatment with 200 mmol·L-1 mannitol; B: 200 mmol·L-1甘露醇处理后 200 mmol·L-1 mannitol treatment; C: 甘露醇处理后的GUS基因表达量 GUS gene expression after mannitol treatment. *表示差异显著(P<0.05),下同。* represent significant difference at P<0.05 with T test, the same below.

图2 转ZjZFN1基因拟南芥PCR鉴定Fig.2 PCR validation of transgenic A. thaliana CK-: 阴性对照 Negative control; CK+: 阳性对照Positive control; M: Marker; 1~11: 转基因植株Transgenic lines.
2.4 干旱胁迫对转基因拟南芥中脯氨酸、丙二醛含量的影响
脯氨酸是渗透调节的重要物质,在干旱胁迫下发挥着重要作用。丙二醛是反映干旱胁迫对植物损害的重要指标,测定转基因和野生型拟南芥叶片中脯氨酸、丙二醛的含量能在一定程度上反映植物的抗旱性[19]。试验结果表明,正常生长条件下转基因植株与WT之间的脯氨酸、丙二醛含量没有显著差异,而干旱处理21 d后WT中脯氨酸的含量比正常生长条件下的增加了107%,增加的含量分别比ZFN-2、 ZFN-17的多67%、131%(图4A);而干旱处理21 d后的WT中增加的丙二醛含量比转基因拟南芥ZFN-2、ZFN-17少了20%、56%(图4B)。
2.5 干旱胁迫下相关抗性基因的表达分析
为了探究ZjZFN1基因在转基因拟南芥中的表达水平,对转基因拟南芥植株 ZFN-2、ZFN-17和野生型拟南芥中的ZjZFN1基因进行qRT-PCR分析,表达分析表明转基因拟南芥中的ZjZFN1的表达水平显著高于野生型拟南芥中的表达水平(图5)。为了进一步探究转ZjZFN1基因是否会影响转基因拟南芥中其他抗性相关基因的表达,对APX、POD、SOD、P5CS、LEA这5种基因进行了qRT-PCR分析。结果显示,干旱处理21 d后ZFN-2、ZFN-17中APX的表达水平均比WT中的提高了59% 左右(图5);而POD的表达水平比WT中的分别下降了54%、60%(图5);SOD、P5CS的表达水平明显下降(图5),SOD的表达水平比WT中的分别下降了46%、59%,P5CS的表达水平比WT中的分别下降了74%、75%;LEA的表达水平比WT中的表达水平分别下降了13%、25%(图5)。
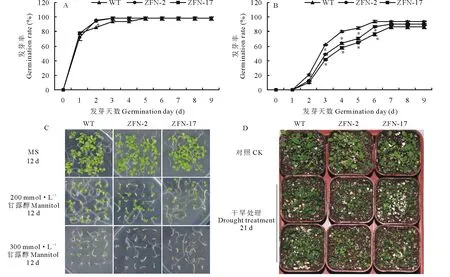
图3 干旱处理下野生型和转基因拟南芥Fig.3 Drought treatment of wide type (WT) and transgenic A. thaliana A: MS上的发芽率 Germination rate on MS; B: 在含有200 mmol·L-1甘露醇的MS上的发芽率 Germination rate on MS containing 200 mmol·L-1 mannitol; C: 在含有不同浓度甘露醇的MS上的发芽情况 Germination on MS containing different concentrations of mannitol; D: 干旱处理 Drought treatment.

图4 干旱处理后野生型和转基因拟南芥Fig.4 Wild-type and transgenic Arabidopsis seedlings after drought treatment 不同小写字母代表单因素方差分析数据差异显著(P<0.05)。下同。The different lowercase letters represent significant differences in one-way ANOVA data (P<0.05). The same below.

图5 干旱处理下转基因拟南芥相关抗性基因的表达Fig.5 Expression of related resistance genes under drought treatment in transgenic Arabidopsis
3 讨论
锌指蛋白是植物中一类重要的转录因子,目前已经在水稻、拟南芥、棉花(Gossypiumhirsutum)、大豆(Glycinemax)等植物中相继被研究[20]。本实验从日本结缕草中分离出一种新的C2H2型锌指蛋白ZjZFN1,旨在更好地了解其功能和潜在的转录调控机制。在前期研究中,我们获得了ZjZFN1基因及其启动子的序列。对转ZjZFN1-Pro::GUS拟南芥植株GUS染色后叶片呈现蓝色,结果表明ZjZFN1的启动子能够驱动GUS基因的表达。对转ZjZFN1-Pro::GUS拟南芥进行甘露醇处理,荧光定量分析表明甘露醇处理后GUS基因的表达量显著增加,这表明ZjZFN1的启动子上可能存在着干旱响应元件。同时,对甘露醇处理后的转基因植株进行GUS染色,结果表明处理后的染色强于未处理之前的,与荧光定量结果相符。在此基础上,通过花序侵染法侵染拟南芥,利用草铵膦筛选拟南芥种子和基因组PCR验证获得了转基因拟南芥。目前,C2H2型锌指蛋白已经从不同的物种中相继被克隆、转化并进行了相关的功能研究,本实验通过对日本结缕草ZjZFN1转化拟南芥探究C2H2锌指蛋白ZjZFN1的功能。
研究发现,不同物种中C2H2型锌指蛋白在干旱胁迫响应中起着重要作用,多数植物的C2H2型锌指蛋白转录因子在干旱胁迫中起正调控的作用,提高植物的抗旱性[21-22]。本研究中,在含有甘露醇的MS培养基中生长12 d后,WT的发芽率高于转基因植株的;干旱处理21 d后,转基因拟南芥相比WT植株矮小、叶片萎蔫。这表明ZjZFN1转录因子可能在干旱胁迫的种子萌发过程中起到负调控的作用,过表达ZjZFN1可能降低了植物的抗旱性,这与玉米(Zeamays)中ZmAN13的功能类似[23-24]。研究表明,ZmAN14与ZmAN13具有很高的同源性,而ZmAN14可以提高植物的耐旱性[23],因此不同的锌指蛋白转录因子在植物的生长发育过程中发挥的功能不同。
为了进一步探究干旱胁迫下ZjZFN1对植物生理功能的影响,本实验对干旱处理后转基因拟南芥ZFN-2、ZFN-17株系中脯氨酸、丙二醛的含量进行了测定。脯氨酸是植物体内的一种重要的渗透调节物质,在干旱胁迫时在植物体内会积累大量的游离脯氨酸[25-27]。丙二醛是植物细胞膜脂过氧化的产物,其含量可表示细胞膜脂的过氧化程度,在一定程度上反映了干旱胁迫对植物造成的损害[19,25]。两者均可作为植物抗旱性的重要指标。实验结果表明,干旱处理后WT、ZFN-2、ZFN-17中的脯氨酸含量都有所升高,而WT中增加的脯氨酸含量高于ZFN-2、ZFN-17中增加的,表明WT的渗透调节功能强于转基因植物。干旱处理后WT、ZFN-2、ZFN-17中的丙二醛含量也都有所升高,而ZFN-2、ZFN-17中增加的丙二醛含量均高于WT中增加的,说明转基因植株中的细胞膜脂损害程度高于WT。干旱处理后植物中脯氨酸、丙二醛含量的变化进一步说明了ZjZFN1在干旱胁迫中可能起负调控的作用。
在植物对环境胁迫的反应中,转录调控起着主导作用[11]。在干旱胁迫下,植物使用活性氧(Reactive oxygen species,ROS)清除酶(如APX,SOD和POD)以消除ROS积累[28-29]。干旱处理后在转基因植株ZFN-2、ZFN-17中APX的表达水平比WT中的表达水平高;而SOD、POD的表达情况与APX的相反,SOD、POD的转录水平的提高反映了ZjZFN1过表达株系中ROS清除能力的增强,这与ZFP179报道的一致[30]。转基因植株中APX与SOD、POD表达水平的不同表明ZjZFN1过表达对不同活性氧清除酶的影响不同。P5CS能控制植物体内脯氨酸的生物合成以增强渗透胁迫下不同酶的活性,被认为是一种重要的抗逆相关基因[31-32]。干旱处理后转基因植株ZFN-2、ZFN-17中的P5CS的表达水平低于WT中的表达水平,表明转基因拟南芥在干旱胁迫中的耐受性降低。LEA是一种逆境胁迫响应蛋白[33-34],在干旱胁迫后转基因植株ZFN-2、ZFN-17中LEA的表达水平低于WT中的表达水平。通过对这几种抗旱相关基因的表达水平的分析,ZjZFN1的过表达影响抗旱相关基因的表达,从而减弱了转基因株系的抗旱性。后期将进行下一步研究,获得RNAi干扰体系或通过CRISPR技术将该盐敏感型基因敲除,从而提高植株的抗旱性。
4 结论
本研究成功将ZjZFN1基因及其启动子转化拟南芥,获得转基因植株。GUS化学组织染色表明ZjZFN1的启动子能够驱动GUS基因的表达,在甘露醇处理下GUS基因的表达水平显著升高。在干旱胁迫下,野生型拟南芥先于转基因拟南芥ZFN-2、ZFN-17株系发芽;干旱处理21 d后,转基因株系相比野生型拟南芥植株矮小、萎蔫,脯氨酸含量低于野生型拟南芥,而丙二醛含量高于野生型拟南芥。干旱处理下,转基因拟南芥中POD、SOD、P5SC、LED的表达水平降低,而APX的表达水平升高。本研究表明ZjZFN1可能减弱植物的抗旱性,为进一步深入研究其功能奠定了基础。
