柳枝稷分蘖发育调控的生理生化机制研究
2019-04-16郑爱泉徐开杰程亭亭奚亚军孙风丽
郑爱泉, 徐开杰, 程亭亭, 张 超, 奚亚军*, 孙风丽*
(1. 西北农林科技大学农学院, 陕西 杨凌 712100; 2. 杨凌职业技术学院, 陕西 杨凌 712100; 3. 中国农业科学院棉花研究所, 河南 安阳 455000)
禾本科植物分蘖是分枝的一种特殊形式,是产量的重要因素之一。其形成包括分蘖芽的起始和伸长两个过程。研究分蘖芽伸长的调控因素可以为生产中提高分蘖数目和生物产量提供指导。分蘖芽伸长是非常复杂的发育过程,受到遗传因素和环境因素共同调控,植物激素是遗传调控和环境调控分蘖芽伸长的直接调节物质[1]。直接参与调控腋芽伸长的内源激素主要包括生长素、细胞分裂素和独脚金内酯,其他激素如脱落酸、油菜素内酯和赤霉素等也通过激素之间的相互作用参与其中。环境因素包括营养成分(氮磷钾等)、水分和光周期等,通过与不同激素的相互作用参与调控分蘖芽/腋芽伸长[2-3]。
生长素(3-吲哚乙酸,IAA)是研究最早的调控植物株型的植物激素之一。植物地上部分中,生长素在主茎茎尖和幼嫩叶片内合成,借助于PIN、AUX/LAX和MDR/PGP家族蛋白通过维管束向茎基部进行极性运输,导致生长素在植株不同部位梯度分布,启动下游信号转导,对胚的发育、器官形成、侧枝发育、侧根产生、维管束发育及植物的向性等多方面具有重要作用[4-5]。细胞分裂素(Cytokinin,CTKs)对腋芽发育具有重要调节作用,可能作为生长素的第二信使直接促进腋芽的生长[6]。CTK含量上升可导致植物在发育后期呈现高度分枝的表型[7]。独脚金内酯(Strigolactones,SLs)是最近被确认的一种新的植物激素,它是一种产生于植物根部的类胡萝卜素衍生物,具有刺激寄生植物种子的萌发和促进丛枝菌根真菌菌丝分枝的作用,可以沿茎干向上运输,与生长素和细胞分裂素一起直接或间接抑制植物分枝[8]。独脚金内酯和生长素、细胞分裂素等其他激素以及环境因素等存在相互作用共同调控腋芽的伸长。其中独脚金内酯与生长素的相互作用是目前研究较多的领域。独脚金内酯与生长素在调控植物腋芽伸长发育中存在密切、复杂的相互作用,拟南芥max突变体中生长素对腋芽伸长的抑制作用降低[9];独脚金内酯可以调节生长素的运输进而影响生长素对植物生长发育的调控作用[10];生长素参与调节SLs的合成[9,11-13]。生长素和独脚金内酯相互作用非常复杂,存在其他激素、环境等调节因素的参与。环境因素中的营养因素如氮(N)和磷(P)等影响植物腋芽的发育,研究发现氮和磷可能通过植物激素调节腋芽的发育,植物在低磷的环境下通过提高生长素受体TIR1的表达增加对生长素的敏感性[14];同时低磷低氮影响植物的SLs合成及运输[15-16];生长素和独脚金内酯信号途径是氮调控植物分枝发育所必需的[17]。
柳枝稷是一种原产于美洲的多年生C4植物,适应性强,生物量巨大,环境适应性强,不占用作物耕地面积,是理想的能源作物之一[18-19]。能源作物年生物产量是重要的生产指标,分蘖数目直接影响柳枝稷的生物产量。分蘖芽分化形成后是否伸长决定了其是否发育成分蘖,柳枝稷分蘖芽的伸长决定了柳枝稷的分蘖数目和生物产量。本研究对一个EMS诱变获得的少分蘖突变体进行生理、生化和分子生物学研究,探索柳枝稷分蘖伸长调控的调控机理,为提高柳枝稷生物量提供分子基础和理论指导。
1 材料与方法
1.1 实验材料
柳枝稷品种Alamo经EMS(甲基磺酸乙酯)诱变所得少分蘖突变体lt,由本实验室创制保存,该突变体材料利用无性繁殖方式保存。生长条件为光照16 h黑暗8 h、光照强度为12 000 lx。
瓜列当种子由西北农林科技大学马永清研究员课题组惠赠。
1.2 实验方法
1.2.1重力反应测定 通过无性繁殖得到的两突变体小苗,在温室小盆中培养15天后,旋转90度,每两天测量地上部分向上弯曲的角度。生长素抑制剂处理选择2,3,5-三碘苯甲酸溶解(TIBA)在丙酮中,随后加入蒸馏水至终浓度为10 μM。每2天加一次相同丙酮含量的自来水作为对照。
1.2.2去顶处理 大田种植的柳枝稷生长至主茎花序伸出时,去掉第一节,7天后测量腋芽伸长的长度。
1.2.3激素测定 激素测定实验由南京钟鼎生物技术有限公司测定,方法如下:
(1)激素提取
准确称量约0.5 g新鲜植物样品,于液氮中研磨至粉碎;向粉末中加入5 ml异丙醇/盐酸提取缓冲液,4℃震荡 30 min;加入 10 ml二氯甲烷,4℃震荡 30 min;4℃,13 000 rpm离心5 min,取下层有机相;避光,以氮气吹干有机相,以 400 μl甲醇(0.1%甲酸)溶解;过 0.22 μm滤膜,进HPLC-MS/MS检测。
(2)液质检测
用色谱柱(Poroshell 120 SB-C18反相色谱柱(2.1×150,2.7 um);柱温:30℃)吸附,流动相:A:B=(甲醇/0.1%甲酸)∶(水/0.1%甲酸);洗脱梯度:0~2 min,A=20 %;2~4 min,A递增至50 %;4~10 min,A递增至80 %;10~11 min,A=80%;11.1 min,A递减至 20 %;11.1~15 min,A=20 %,进样体积为2 μl。质谱条件:气帘气:15 psi;喷雾电压:4 500 v;雾化气压力:65 psi;辅助气压力:70 psi;雾化温度:400℃
1.2.4瓜列当种子发芽 瓜列当种子用10%次氯酸钠溶液灭菌3 min,无菌水洗涤3~4次,用70%乙醇洗涤1 min,无菌水洗涤3~5次,超净台晾干后待用。将瓜列当种子放置在加水的滤纸上在25℃的培养箱中暗培养3d。取柳枝稷根0.5 g用液氮研磨,加1 ml甲醇超声处理30 min,离心取上清液。稀释10倍后处理预培养的瓜列当种子,3 d后统计发芽率。
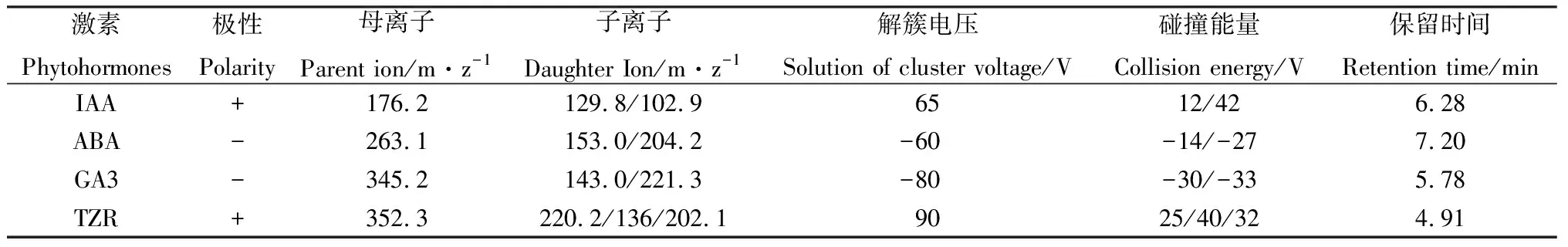
表1 不同激素的检测条件Table 1 Tests of different conditions of the corresponding hormones
1.2.5RNA提取、反转录与定量PCR 取新发出的柳枝稷分蘖使用Trizol方法进行总RNA提取,使用Takara反转录试剂盒进行cDNA第一链合成。根据Primer 5.0设计定量PCR引物(表2),按照荧光定量PCR试剂盒(Takara)使用说明,使用ABI Q7定量PCR仪进行实验,利用其指定的软件进行数据分析。

表2 定量PCR引物列表Table 2 The primers used in qPCR
1.2.6数据统计 文中测量数据利用Excel 2010做图表,利用LDS法对各试验数据进行方差分析。
2 结果与分析
2.1 lt突变体分蘖能力减弱
与对照相比,lt突变体具有较弱的分蘖能力。突变当代lt突变体可产生有2个分蘖而对照含有36个分蘖。柳枝稷是自交不亲和植物,突变体采用无性繁殖。无性繁殖后代的第一代lt突变体和对照相比分蘖能力依然较弱(图1)。
2.2 去顶后lt突变体腋芽伸长速度减缓
柳枝稷茎节的腋芽一般处于休眠状态,去掉茎尖顶端分生组织后,生长素对茎节腋芽的抑制作用解除,腋芽开始伸长。lt突变体去顶后所有腋芽的伸长速度与对照相比明显降低(P<0.01)(图2)。对照中从顶部向下的第一腋芽和第二腋芽的去顶后的伸长速度无明显差异(P>0.05),而lt突变体中第一腋芽伸长速度明显高于第二腋芽伸长速度(P<0.05)(图2)。这些结果表明lt突变体去顶后腋芽的伸长速度受到抑制。

图1 突变体lt分蘖能力下降Fig.1 The tillering ability of the lt mutant was decreased注:(A)和(B)分别为对照ht和突变体lt刈割第二代表型,Bar=10 cm;(C)分别统计对照ht和lt突变体刈割后代的分蘖数;误差线表示SEM;n=5. V0表示种子长出的当代;V1表示第一次刈割后长出的后代Note:(A) The phenotype of V1 from ht and lt. Bars=10 cm. (B) Statistics tiller number of vegetable generations from ht and lt separately. Error bars represent SEM;n=5. V0 means the seedling grow from the isolated buds. V1 means the second generation after cutting
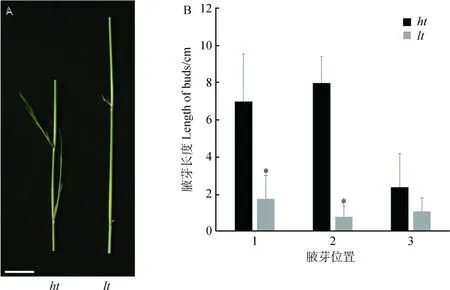
图2 去顶后突变体lt腋芽伸长受抑制Fig.2 The speed of axillary bud outgrowth in lt were inhibited after decapitation注:A去顶后腋芽的伸长B统计结果,1、2、3代表腋芽的从上往下的顺序Note:(A) Photograph of auxillary bud outgrowth in ht and lt after decapitation. (Left:ht;Right:lt) (B) Statistical analysis of length of auxillary bud outgrowth at different shoot nodes in ht and lt
2.3 lt突变体的重力反应减弱
lt突变体地上部分的重力反应测定发现,lt突变体的重力反应较对照弱(P<0.01)(图3A)。用生长素运输抑制剂TIBA处理,二者的重力反应均变弱,但两者之间的差异并未减少(P>0.05)(图3B)。这些结果表明生长素运输缺陷并不是导致lt突变体与对照向重力性反应差异的主要因素。

图3 ht和lt茎的向重力性Fig.3 Shoot gravitropism in switchgrass ht and lt注:A.重力刺激下ht和lt的表型(左:ht;右:lt)箭头指向代表重力方向,标尺为5厘米。B. 统计分析TIBA处理过和未处理过ht和lt茎的弯曲度.误差线表示SEM,n=10,P<0.01 (ANOVA)Note:A.Photographs of ht and lt upon gravistimulation (left:ht;Right: lt) white arrow:direction of gravity. B.Statistical analysis of shoot curvature in ht and lt seedlings with or without TIBA treatment. Error bars represent SEM;n = 10,P<0.01 (ANOVA)
2.4 lt突变体中内源植物激素IAA和ABA含量增加
为了研究促进分蘖突变体形成的机制,用GC/MS方法测量了两个突变体中赤霉素、脱落酸、生长素和细胞分裂素的含量。lt突变体中生长素和脱落酸的含量相比对照明显升高(P<0.05),赤霉素和细胞分裂素的含量与对照无明显区别(P>0.05)(图4)。由此推测lt中分蘖芽生长受抑制可能与生长素和ABA的高含量有关。
2.5 lt突变体独脚金内酯(SLs)含量增加
独脚金内酯(SLs)最近被确定为激素抑制分枝[20-21]。SL可以刺激列当种子萌发,植株根系提取物刺激列当种子发芽可以间接反映独脚金内酯含量。结果表明lt突变体的根系提取物刺激列当种子发芽率高于ht对照,推测lt突变体中独脚金内酯的含量高于对照,高含量的独脚金内酯导致lt的分蘖发育受到抑制(图5 A-E)。
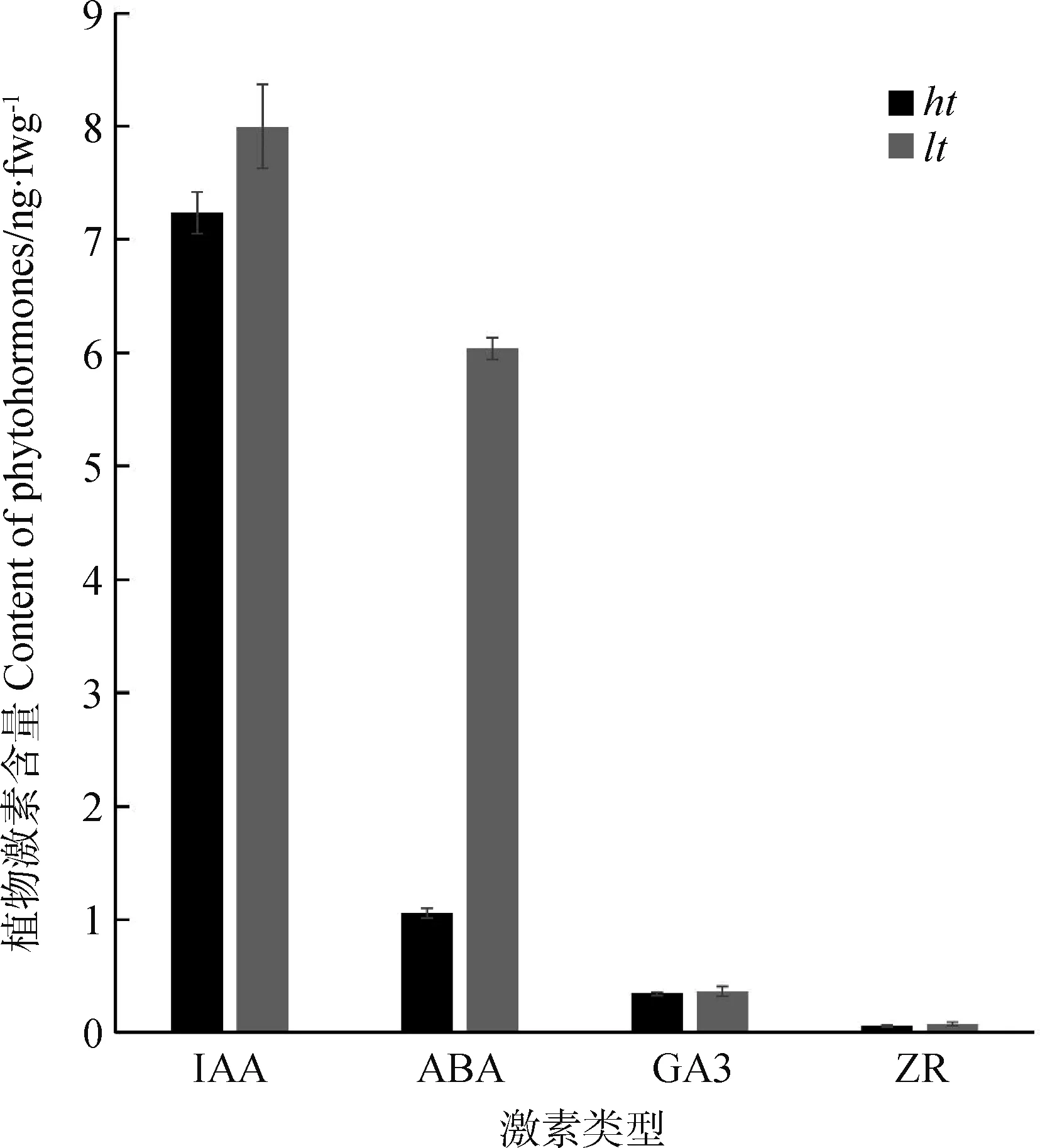
图4 突变体lt的植物激素含量变化Fig.4 Hormone levels in ht and lt mutant注:误差线表示± SDs;n = 3. *代表ht和lt之间存在差异显著(P<0.05,ANOVA),**代表ht和lt之间存在极显著差异(P<0.05,ANOVA)。Note:Error bars represent means±SDs;n = 3. *There were a significant difference between ht and lt (P<0.05,ANOVA). **There were a highly significant difference between ht and lt(P<0.01,ANOVA)
独脚金内酯合成及信号转导途径的关键基因已经较为明确,水稻中D10、D17和D27参与类胡萝合成SLs的过程,D14作为SLs的受体与D3、D53组成复合体,传递SLs的信号,对下游基因进行调控进而影响分蘖的发育。检测ht和lt突变体中这些基因的表达量发现D3、D10、D14、D17和D27等在lt突变体中高于ht对照(P<0.01)。SLs信号途径的抑制子D53在lt突变体中表达水平略低于ht对照(P<0.05)(图5 F)。结果表明,lt突变体中SLs的合成和信号相对于对照增强,独脚金内酯合成的增加是造成lt突变体分蘖数目减少的重要因素。

图5 ht和lt中独脚金内酯含量分析及相关基因的检测Fig.5 Analysis of strigolactones content and expression level of related genes in ht and lt mutant.注:A-E为不同物质刺激瓜列当种子萌发。A.水;B. GR24;C. ht根际分泌物;D. lt根际分泌物;E. 统计分析;误差线表示± SDs;n=3。ht和lt呈极显著差异(P<0.01,ANOVA). 标尺为1 mm。F. 独脚金内酯途径相关基因在ht和lt中的表达量变化(Actin基因为内参)。**代表差异极显著(P<0.01);*代表差异显著性(P<0.05)Note:A. Water;B. GR24;C. Combined root exudates from ht plant;D. Combined root exudates from the lt mutant;E. Statistical analysis of germination stimulation on O. aegyptiaca seeds. Error bars represent means±SDs;n=3. There were a significant difference between ht and lt (P<0.01,ANOVA). Bars=1 mm;F. Relative expression level of genes involved in SLs. The ACTIN gene was the reference gene. **There were a highly significant difference between ht and lt (P<0.01,ANOVA). *There were a significant difference between ht and lt (P<0.05,ANOVA)
3 讨论
分蘖芽伸长是由植物激素和环境之间的相互作用调控的,生长素由枝条顶端分生组织产生,通过极性运输运送到基部,抑制腋芽的生长,而CK被向顶部运输,同时促进腋芽生长,SLs同样向顶运输,但抑制腋芽生长[10]。ABA可以通过影响生长素合成和运输来影响腋芽生长[22]。这些植物激素通过形成一个网络来调控植物内部生长。柳枝稷的少分蘖突变体lt的独脚金内酯、ABA和生长素含量均有所升高。定量PCR结果显示参与SLs生物合成和信号途径的基因表达水平存在差异。因此,柳枝稷分蘖芽的形成可能受上述植物激素的综合调控。
水稻的SL信号途径突变体能够产生大量分蘖并且能够增加突变体的向重力性反应。SLs水平降低会导致较低一侧的茎基部生长素合成增加,从而增强了茎部的向重力性[23]。本研究发现,lt突变体中可能由于SLs的含量较高,导致SL信号途径相关基因表达水平较高从而抑制分蘖芽的发育(图5)。同时lt茎部的向重力反应较弱(图3),可能是由于lt突变体的SLs水平升高导致较低一侧的茎基部生长素合成减少降低茎部的向重力性。但是lt中生长素的含量增加(图4),lt突变体植株地上部分的向重力性反应减弱可能是由于生长素分布异常导致的,但是生长素运输抑制剂TIBA处理后并不能减少lt与ht对重力响应的差异,有待验证生长素运输相关的其他途径。同时本研究中SLs含量检测是用提取物刺激列当种子发芽进行间接测量的,需要更直接的方法检测来进一步研究柳枝稷中SLs对分蘖的调控机制。
多年生禾本科植物柳枝稷能够从位于茎节、根冠及根茎产生分蘖芽,在本研究中,突变体中分蘖芽的形成和伸长均受到抑制。柳枝稷为自交不亲和植物,所以试验中的突变体是种子通过EMS诱变产生后通过无性繁殖保存的,突变体与对照之间可能会有遗传上的差异,即ht并不是严格意义的对照。同时我们发现突变体lt与对照ht之间的存在其他的差别,包括分蘖高度和茎秆直径等方面,这可能是由于基因的一因多效造成的,也有可能是存在遗传背景的差异,后期将在连续的生长期内对突变体调控分蘖的特殊机制做进一步研究。
4 结论
本研究对柳枝稷少分蘖突变体lt进行生理、生化和分子生物学研究,分析少分蘖突变体分蘖数目减少的原因。结果表明,突变体中的生长素、脱落酸和独脚金内酯等植物激素含量的增加可能是其分蘖芽伸长发育受抑制的重要因素。本研究可为生产中提高柳枝稷分蘖数增加生物产量提供理论指导。
