猫外膝体中脑源性神经营养因子及其受体表达的年龄相关性变化
2019-03-11童传旺
童传旺
(芜湖职业技术学院,安徽芜湖241000)
在正常衰老过程中,视觉能力会出现进行性衰退,表现为视觉对比敏锐度下降、视锐度减弱、对物体朝向和运动方向辨别能力的障碍以及对视觉信息分析速度减慢等[1-2],有关老年性视觉功能衰退的神经机制尚处于探索阶段[3]。一方面,老年性视觉功能衰退主要与视觉通路中哪些层次的神经可塑性变化有关仍处于争论之中[1,2,4-5]。尽管一些电生理研究结果显示,外膝体(lateral geniculate nucleus,LGN)神经元对视觉刺激的反应特性没有明显的年龄相关性变化,衰老可能主要影响视觉皮层特别是高级视觉皮层神经元的功能[1,4-6],但这并不能排除衰老过程中LGN可能发生的形态学和神经递质系统的改变[7],这些改变可通过突触联系影响视皮层对视觉刺激的反应特性。另一方面,老化引起视觉中枢神经元功能衰退的细胞分子机制尚不清楚。最近的研究结果显示,衰老伴随着视觉中枢内抑制性神经递质(如GABA)表达减少[8],神经抑制作用减弱,这可能是导致视觉皮层神经元对视觉刺激反应的敏感性和选择性下降的重要原因[9],然而,引起抑制性神经递质系统活动降低的信号传递和分子机制却知之甚少。
近些年对神经营养因子的研究发现,脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)通过其受体介导的信号转导在神经元的发育、存活、损伤修复和神经可塑性等方面发挥着重要作用[10-12]。有报道显示,BDNF信号系统可参与皮层内活动依赖的抑制行调节,并促进GABA的合成和运输[13-15]。是否因衰老过程中BDNF及其受体表达下降,从而引起抑制性神经递质GABA分泌减少?目前尚无充足的试验证据,尽管有少数研究报道显示,脑内BDNF及其受体的表达出现了年龄相关性变化[16]。
猫作为研究视觉系统功能的模型动物,因其视觉系统进化较为高级且视觉组织结构与系统功能与人类较为相似。本研究选取外膝体作为视觉功能研究层次,基于外膝体在第一视觉通路中具有视觉信息初级整合编码的“中继站”作用,希望通过对外膝体开展相关研究,揭示外膝体水平上的有关视觉通路相关调节机制的研究资料。我们以前的研究发现,猫外膝体(LGN)内抑制性神经递质GABA的表达增龄性显著降低[17],这是否与BDNF及其受体的表达下调有关尚不清楚。为进一步探讨该问题,本研究以青、老年猫为研究对象,采用免疫组织化学方法研究外膝体中BDNF及其高亲和力的酪氨酸激酶B受体(tyrosine kinase receptor,TrkB)表达以及低亲和力受体蛋白(P75NTR)的年龄差异,为视觉功能衰老的细胞、分子机理的研究提供思路。
1 材料和方法
1.1 试剂材料及试验动物
本研究选取2.0~3.5 kg体重猫为试验对象,试验前对试验对象行眼部疾病检查确保其视觉功能正常而消除干扰。选定青年组猫(1-3龄)与老年组(10-13龄)各4只进行试验编组。施行盐酸氯胺酮注射麻醉达足够深度后(盐酸氯胺酮按体重40 mg/kg计),立即采取开胸操作,并向心脏内灌注生理盐水(0.9%),待肝脏发白后灌注预固定液(戊二醛2.5%+福尔马林10%+磷酸缓冲液pH7.2~7.4,0.1 mol/L,按体重200 mL/kg计)。开颅操作切取脑干部分继续固定2 h。切取外膝体静置于固定液(戊二醛2.5%+福尔马林10%+蔗糖30%),直至脑组织块完全固定沉入底部。制作厚20μm冰冻冠状连续切片,隔5取4法选取切片编组(单只猫抽取10组),每组4片切片对应BDNF、TrkB、P75NTR免疫染色组及空白对照组。
1.2 免疫组织化学染色
室温下滴注3% H2O2孵育、灭活内源性过氧化物酶冰冻切片15 min;纯水淋洗后,室温下滴注PBS溶液(含TritonX-100:0.3%)孵育20 min;室温下滴注小牛血清蛋白(5%)至完全封覆特异反应部位孵育20 min;倾尽血清,滴注经1∶100稀释的一抗(分别为兔抗鼠BDNF及兔抗人TrkB、P75NTR多克隆抗体稀释液)4℃恒温孵育36 h;以PBS淋洗(5 min为1次,连续3次);滴注二抗(羊抗兔IgG工作液)进行反应标记,室温下孵育20 min;以PBS淋洗(5 min为1次,连续3次);滴注三抗SABC(链霉素过氧化物酶),室温下孵育20 min;以PBS淋洗(5 min为1次,连续3次);DAB染色显色,切片经常规梯度浓度酒精脱水操作后,二甲苯透明,中性树胶封片并作镜下检查。空白对照切片组,一抗代之以PBS液,余同处理。试验试剂盒、DAB购自博士德生物公司。
1.3 观察记录形态学数据及分析处理
随机抽取青年、老年组中BDNF、TrkB、P75NTR各20片染色切片进行显微观察,并采集形态信息。观察用显微镜型号为Olympus BX-51,图像采集、分析信息软件采用Image Pro Express 6.0软件完成。主要观察及采集信息指标包括:BDNF、TrkB、P75NTR免疫阳性细胞在猫外膝体各层的表达情况,以及免疫阳性反应强度(以光吸收度均值IOD为参照,反应强度越高,IOD值越大)。
低倍镜下(40×)分别获取BDNF、TrkB、P75NTR染色切片图像,较高倍镜下(100×)确定外膝体A、A1、C层结构,再至高倍镜(400×)下观察计数。以“50μm×50μm”标尺确定视野大小规格,各层视野数随机选定10个。各视野内统计BDNF、TrkB、P75NTR免疫阳性细胞数,染色细胞胞体含有明显的免疫阳性物质且具有细胞核为识别计数标准。经各视野内统计BDNF、TrkB、P75NTR免疫阳性细胞数,多次观察值取均值后结合视野大小转换为细胞密度(cells/mm2)记录。免疫阳性反应强度IOD值测量以图像采集软件对切片随机选取视野(每张切片20个)测量记录。采集数据以“平均值±标准差”(mean±SD)形式记录。分析统计由SPSS 13.0软件进行,统计方法为ANOVA显著性检验及相关性检测。
2 结果与分析
2.1 BDNF及TrkB、P75NTR阳性细胞形态学信息统计分析
切片观察发现,猫外膝体中各层中BDNF及TrkB、P75NTR阳性细胞及纤维均有分布,胞体多呈棕黄或黄褐色。低倍镜下可见外膝体A、A1层免疫阳性细胞分布较C层密集。高倍镜下可见着色较深的呈卵圆形或不规则形阳性细胞胞体,且阳性细胞的突起结构(神经元轴、树突及胶质细胞突起)中亦可见免疫阳性物质分布(见图1)。
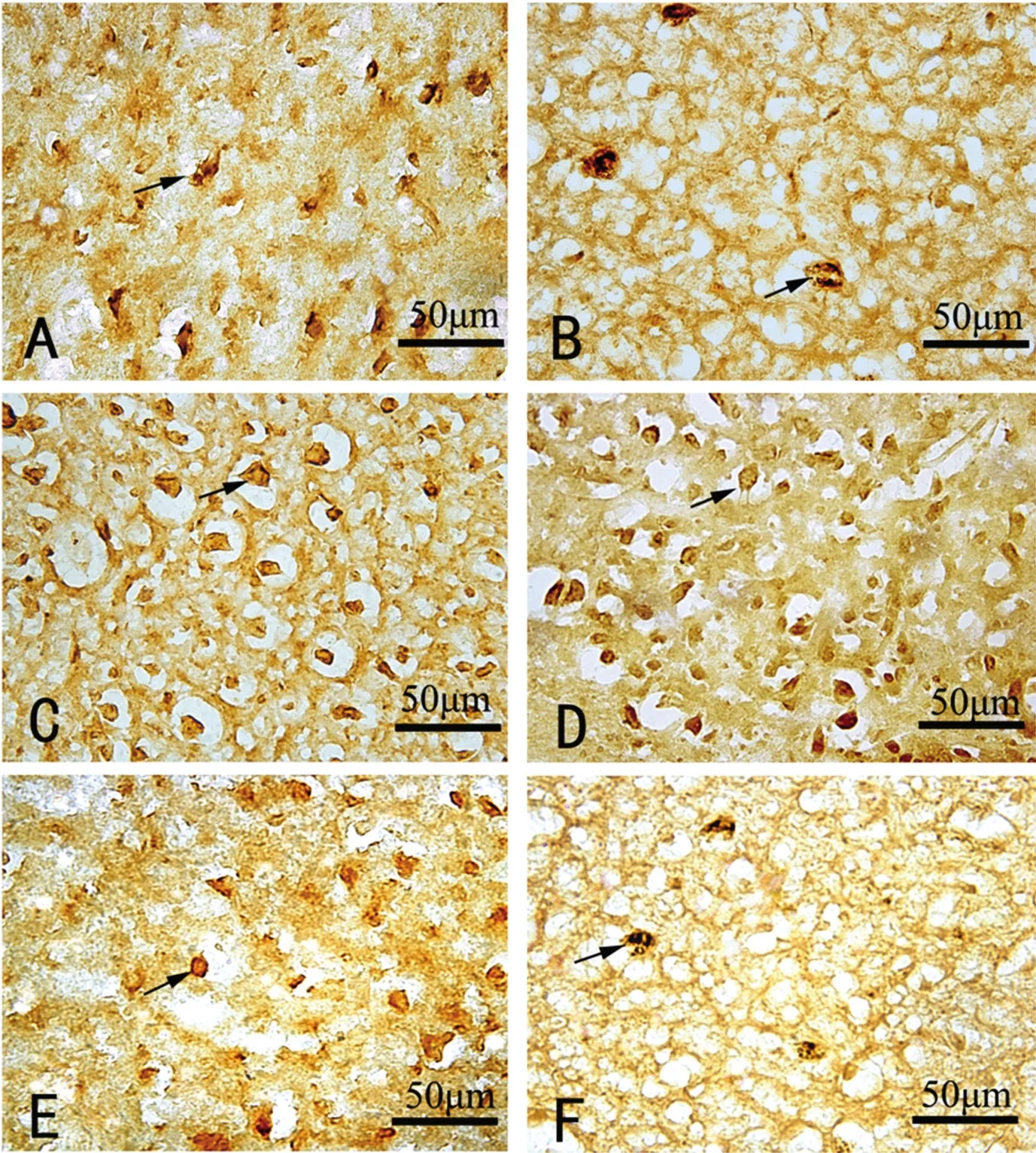
图1 猫外膝体A层中免疫阳性细胞(箭头指及)及纤维Figure 1 Show immunoreactive cells(arrows)and fibers in the LGN(lamina A)of cats
青年、老年组内外膝体不同层次间数据比较检验结果显示,在青年、老年组猫外膝体中,BDNF阳性细胞的密度表现为A层和A1层分别较C层高[F(1,158)=27.12,P<0.01;F(1,158)=40.06,P<0.01],A层 与A1层相比无显著差异[F(1,158)=1.33,P=0.25]。青年、老年组间数据比较结果显示,老年猫外膝体中BDNF免疫阳性细胞密度较青年猫显著降低[F(1,158)=125.39,P<0.01],且各年龄组内个体间无显著差异[F(3,236)=1.11,P=0.35]。青年、老年组内外膝体A层和A1层中的TrkB及P75NTR阳性细胞密度亦较C层的高[TrkB组:F(1,158)=28.16,P<0.01;F(1,158)=29.92,P<0.01。P75NTR组:F(1,158)=44.14,P<0.01;F(1,158)=33.13,P<0.01],A层与A1层相比无显著差异[TrkB组:F(1,158)=0.02,P=0.89。P75NTR组:F(1,158)=1.23,P=0.14]。青年、老年组间数据比较结果显示,老年组TrkB及P75NTR免疫阳性细胞密度均较青年猫的显著减小[TrkB组:F(1,158)=208.54,P<0.01。P75NTR组:F(1,158)=320.58,P<0.01],且各年龄组内个体间无显著差异[TrkB组:F(3,236)=0.22,P=0.88。P75NTR组:F(3,236)=0.94,P=0.25](见表1)。
数据相关性检测分析结果提示:猫外膝体BDNF及TrkB、P75NTR免疫阳性细胞密度变化之间呈现显著相关性(BDNF与TrkB相关系数r=0.87,P<0.01;BDNF与P75NTR相关系数r=0.57,P<0.01;TrkB与P75NTR相关系数r=0.61,P<0.01)。
2.2 BDNF及TrkB、P75NTR受体免疫阳性反应强度
数据分析结果显示:与青年组相比,老年组猫外膝体中BDNF阳性反应强度值显著减小[F(1,78)=18.74,P<0.01],其中A、A1和C层分别下降幅度为26.4%、20.3%和36.3%。老年组TrkB阳性反应强度比青年组显著减小[F(1,78)=28.46,P<0.01],其中A、A1和C层分别下降幅度为25.6%、26.5%和35.3%。老年组P75NTR阳性反应强度比青年组亦显著减小[F(1,78)=39.72,P<0.01],其中A、A1和C层分别下降幅度为21.3%、18.5%和32.1%(见图2)。
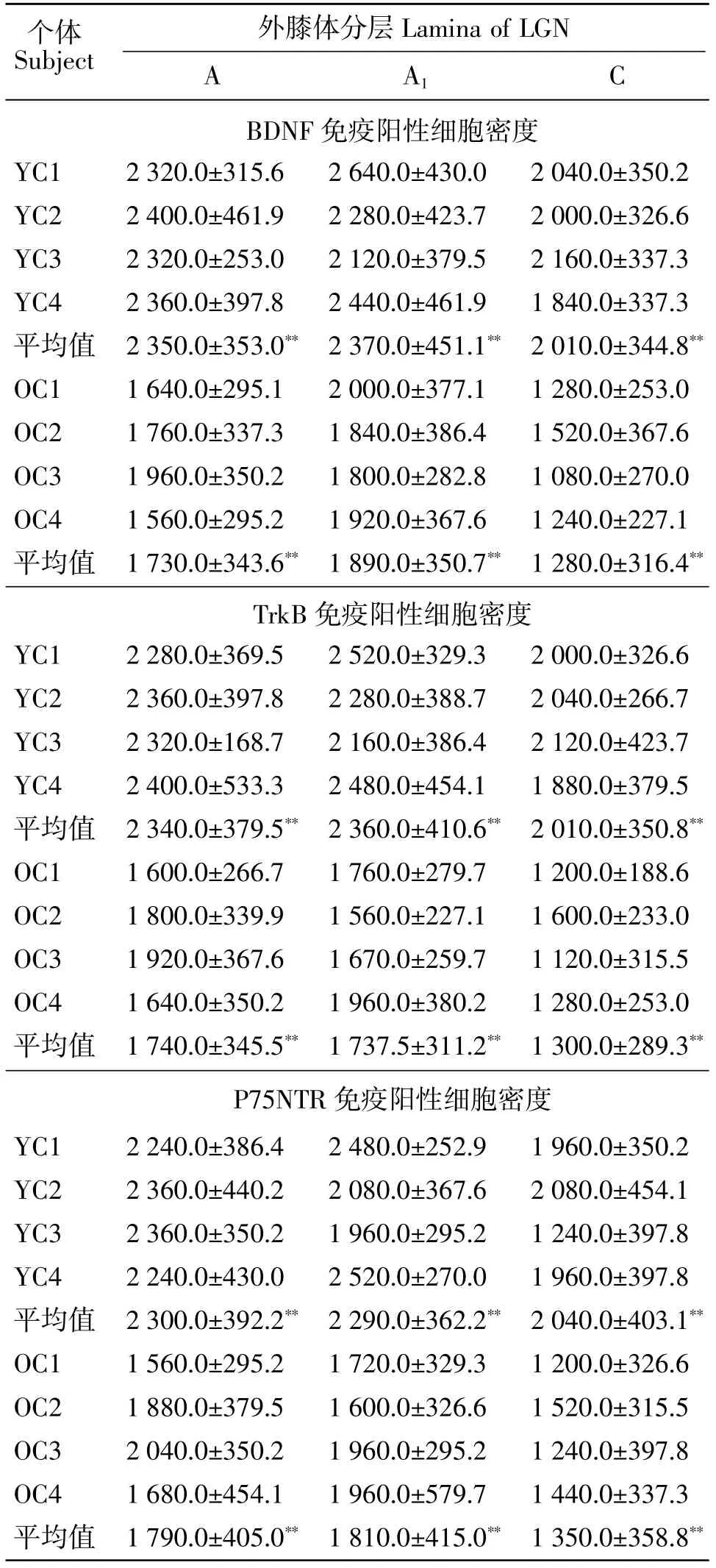
表1 青年组及老年组猫外膝体中BDNF及TrkB、P75NTR免疫阳性细胞密度比较(mean SD)Table 1 A comparison on density of BDNF-IR and TrkB-IR,P75NTR-IR cells in the LGN of young and old cats(mean SD) cells/mm2
3 讨论与结论
高等动物主要依靠视觉功能获得外部信息,衰老引起的视觉功能减退会直接影响生活的质量。探讨老年性视觉能力下降的神经网络和细胞分子机制,有利于通过医学途径延缓衰老对视觉的影响,提高老年人的视觉能力[18]。
在人和高等动物中,视觉信息的加工分视网膜、外膝体和视皮层3个由低到高的层次。近些年的电生理研究结果显示,衰老可能主要改变了视皮层神经元对视觉刺激的反应特性,包括对刺激的方位和运动方向选择性、对比度敏感性以及空间和时间频率的选择性等[19-21],而衰老对外膝体神经元反应特性的影响似乎并不明显[22]。尽管如此,我们并不能排除外膝体在细胞和分子水平的老年性变化。之前的研究发现老年猫外膝体中抑制性神经递质GABA的表达比青年猫的显著减少[17]。神经递质GABA的表达减少可能因为下行通路或其他因素的补偿作为而没有在功能水平表现出来。因此,衰老对外膝体的影响主要表现在细胞水平。
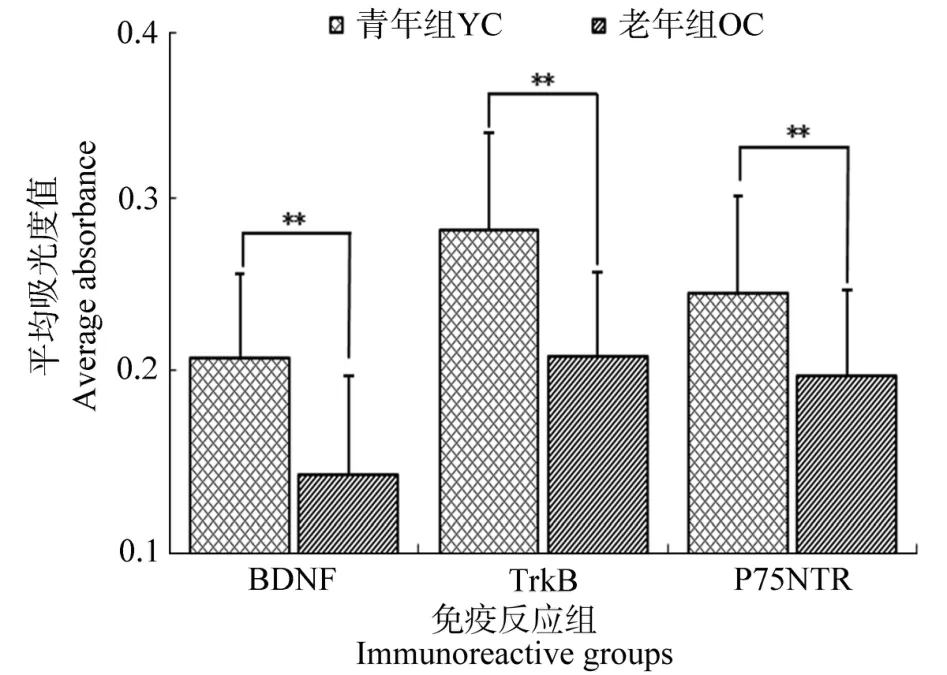
图2 猫外膝体中BDNF、TrkB及P75NTR免疫阳性反应的平均光吸收IOD值Figure 2 Show the mean IOD(Integrated optical density)value of BDNF and TrkB,P75NTR immunoreactivity in the LGN of cats
近些年对神经营养因子的研究结果显示,脑源性神经营养因子BDNF通过其受体介导的信号转导,对神经元的发育、存活及突触可塑性等具有重要作用[23]。BDNF受体主要包括高亲和力的受体Trk和低亲和力的P75NTR受体,其中Trk类受体与BDNF特异性结合直接发挥特定功能,而低亲和力受体P75NTR虽可与各类神经营养因子广泛结合,其并非为神经营养因子发挥功能的必需因子。一般认为P75NTR主要为增强高亲和力受体TrkB与BDNF高亲和结合位点的形成过程,并增强TrkB受体表达的时阶特异性与组织特异性[24]。有相关研究证实,BDNF参与的信号转导对于GABA能神经元的发育、存活和功能表达具有重要的支持作用[25-27],并可能直接参与脑内抑制性神经递质GABA的合成和转运[13-14]。是否老年动物外膝体中GABA表达的下降与BDNF信号转导作用异常引起的,目前尚缺乏充足的试验证据。
本研究结果显示,老年猫和青年组猫外膝体各层神经元密度没有显著差异,表明在衰老过程中外膝体神经元的数量维持相对稳定,这与以前的研究报道结果一致[17]。然而,老年猫外膝体各层中BDNF和TrkB、P75NTR免疫阳性细胞密度比青年猫的显著下降,且BDNF和TrkB、P75NTR的免疫阳性反应强度亦明显减弱。这一结果提示老年组猫中神经元的绝对数量并未发生显著减少,而BDNF和TrkB、P75NTR相关的特异神经元阳性反应呈现出的在数量和强度上的显著减弱提示老年个体BDNF-TrkB调节功能呈现下降趋势。类似的结果也在视觉高级结构初级视皮层BDNF年龄相关性表达变化研究中亦有报告[28]。阿尔茨海默症(Alzheimer disease,AD)患者及老龄大鼠的海马中也曾报道过较一致的观察结果[16,29-30],表明在衰老过程中BDNF与TrkB、P75NTR的表达减少,BDNF-TrkB信号转导活动降低。
总之,本研究结果表明,衰老会引起外膝体各层中BDNF与TrkB、P75NTR受体的表达下调,P75NTR介导的BDNF-TrkB结合过程的活化减弱,BDNF-TrkB信号转导活动减弱可能导致GABA表达下降,这些抑制性神经末梢通过突触前和突触后抑制作用的减弱可能亦会影响视皮层神经元对视觉刺激反应的选择性和敏感性,从而引起老年性视觉能力减退。
