顺铂耐药相关基因的筛选、验证及生物信息学分析∗
2019-02-16王旦旦陈智如饶进军
王旦旦,吕 琳,陈智如,饶进军
(南方医科大学药学院,广州 510515)
2018年全国癌症报告显示,卵巢癌是女性最常见的恶性肿瘤之一,其病死率高居妇科肿瘤之首,其中卵巢上皮癌约占60%[1]。肺癌则是我国各地区男性发病率及病死率最高的恶性肿瘤[2],而肺腺癌是最常见的肺癌类型,约占50%[3]。在晚期卵巢上皮癌及肺腺癌患者中,以顺铂等铂类药物为基础的化学疗法常被用于一线治疗[4],而肿瘤细胞对顺铂产生耐药性是影响其临床治疗效果和患者预后的一大难题[5]。研究表明,肿瘤一旦对顺铂形成耐药性,则第二代的卡铂及第三代的奥沙利铂临床治疗效果也明显减弱。鉴于以上事实,逆转肿瘤细胞对顺铂的耐药性是当前抗肿瘤研究的一个重要方向。肿瘤细胞对顺铂耐药的形成是一个由多种因素共同参与调控的复杂过程[6],其产生的基础是基因的差异表达。因此,通过高通量的基因芯片技术和生物信息学方法全面检测与肿瘤顺铂耐药相关的基因对初步探究顺铂耐药的发生机制具有非常重要的意义[7]。
以往有关顺铂耐药机制的研究往往只针对一种肿瘤细胞,由于卵巢上皮癌和肺腺癌均选用顺铂作为其一线化疗药物且都容易导致耐药,因此本研究基于已发表的人卵巢上皮癌及肺腺癌顺铂耐药基因芯片数据,利用生物信息学方法分析共同差异基因并运用实时荧光定量PCR(qRT-PCR)进行验证,同时分析差异基因表达对患者预后的影响,旨在筛选出在不同肿瘤细胞顺铂耐药形成中共同起作用的关键基因,为临床寻找逆转顺铂耐药和改善患者预后的新靶标提供理论基础,现报道如下。
1 材料与方法
1.1基因芯片数据收集 从基因表达数据库GEO(www.ncbi.nlm.nih.gov/geo)中下载得到ID号为GSE33482的卵巢上皮癌mRNA芯片数据及ID号为GSE108214的肺腺癌mRNA芯片数据,其中GSE33482包含6例人卵巢上皮癌顺铂耐药细胞株A2780/DDP标本和6例人卵巢上皮癌亲本敏感细胞株A2780标本;GSE108214包含5例人肺腺癌顺铂耐药细胞株A549/DDP标本和4例人肺腺癌亲本敏感细胞株A549标本。
1.2方法
1.2.1共同差异基因筛选 将GSE33482和GSE108214中的芯片数据导入R语言(https://www.r-project.org/)软件中,使用affy包对上述两个芯片的原始数据进行过滤和标准化处理,并运用Limma包中的经验贝叶斯方法分别筛选出差异表达基因,筛选阈值设为校正后P<0.05,差异表达倍数|logFC|≥1,然后使用edgeR包绘制差异基因的热图。运用VENNY2.1(http://bioinfogp.cnb.csic.es/tools/venny/index.html)在线工具,分别对在卵巢上皮癌及肺腺癌耐药株中表达均为上调或下调的差异基因取交集得到共同差异基因。下述的生物信息学分析均只对共同差异基因进行分析。
1.2.2共同差异基因富集分析 将共同差异基因分别导入Metascape数据库(http://metascape.org/gp/index.html)中进行GO功能和KEGG通路富集分析(设定参数:Overlap≥3,Enrichment≥1.5)。通过Fisher Exact Test计算P值,以P<0.05为差异有统计学意义。
1.2.3蛋白质相互作用网络分析 运用String10.5(https://string-db.org)构建共同差异基因的蛋白相互作用(protein-protein interaction network,PPI)网络,将最低互作分值设置成高度可信(high confidence=0.7),并将分析结果转入Cytoscape软件中进行可视化处理,然后通过软件中的Network analyzer插件筛选出节点较多的关键基因并构建PPI核心网络。
1.2.4qRT-PCR验证基因表达 分别收集本实验室保存的卵巢上皮癌顺铂耐药细胞株A2780/DDP及亲本敏感株A2780与肺腺癌顺铂耐药细胞株A549/DDP及亲本敏感株A549,运用Trizol提取总RNA。按照反转录试剂盒(PrimeScriptTMRT reagent Kit with gDNA Eraser,Takara)说明书合成cDNA,然后根据SYBR定量PCR试剂盒(TB GreenTMPremix Ex TaqTM,Takara)操作步骤配制反应体系,在实时荧光定量PCR(RT-qPCR)仪(LightCycler96,Roche)上进行反应。选用甘油醛-3-磷酸脱氢酶(GAPDH)基因作为内参基因,各基因引物序列见表1。分别以A2780及A549中各基因的mRNA表达水平为对照,根据2-△△Ct值计算关键差异基因在A2780/DDP及A549/DDP中mRNA的相对表达水平。
1.2.5生存分析 使用Kaplan-Meier Plotter(http://kmplot.com/analysis/)分析关键差异基因的表达对卵巢癌和肺腺癌患者生存率的影响,采用Kaplan-Meier法分析并绘制生存曲线,通过Log-rank检验法比较生存曲线的差异,以P<0.05为差异有统计学意义。
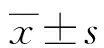
2 结 果
2.1筛选的共同差异基因 在A2780/DDP与A2780之间共筛选得到2 457个在耐药细胞株中表达上调和1 496个表达下调的差异基因;在A549/DDP与A549之间共筛选得到1 718个在耐药株细胞中表达上调和1 512个表达下调的差异基因(校正后P<0.05,|logFC|≥1)。差异基因的热图见图1A、B。分别对在两种耐药细胞株中表达上调和下调的差异基因取交集,最终得到295个表达上调和176个表达下调的共同差异基因,见图1C、D。

表1 qRT-PCR引物序列

A:芯片GSE33482差异基因热图;B:芯片GSE108214差异基因热图;C:表达上调差异基因韦恩图;D:表达下调差异基因韦恩图
图1筛选的共同差异基因热图及韦恩图

表2 共同差异基因的GO功能和KEGG通路富集分析(部分)
2.2共同差异基因的GO功能和KEGG通路富集分析 富集分析结果(P<0.05和FDR<0.05)显示,在耐药株中表达上调的共同差异基因主要富集于损伤修复、辅助性T淋巴细胞17(Th17)细胞分化和磷脂酰肌醇-3-激酶/丝苏氨酸蛋白激酶(PI3K-AKT)信号通路等;表达下调的共同差异基因则主要富集于细胞黏附、轴突导向和细胞因子间受体相互作用等,见表2。
2.3蛋白相互作用网络分析 蛋白互作分析得到包含273个节点的PPI网络。其中,PPI核心网络由相互作用关系较高的10个关键基因RIPK4、JUN、PRKCA、JAK1、HSPA1A、HSPB1、PLCB1、GNG4、SHC1和CXCL1构成。PPI核心网络与其他节点有较多连接,提示上述基因可能起关键作用,见图2。

A:共同差异基因的蛋白互作网络图;B:关键差异基因核心网络图
2.4qRT-PCR检测结果 运用qRT-PCR验证10个关键差异基因的mRNA表达情况。RIPK4、JUN、PRKCA、JAK1、HSPA1A、PLCB1、GNG4、SHC1基因在A2780/DDP及A549/DDP细胞中的mRNA表达水平均分别高于A2780及A549细胞(P<0.05),CXCL1基因在A2780/DDP及A549/DDP细胞中的mRNA表达水平低于A2780及A549细胞 (P<0.05),与芯片结果一致。但HSPB1在A2780/DDP中的mRNA表达水平却低于A2780,与芯片数据不一致,见图3。
2.5生存分析 分析关键差异基因RIPK4、JUN、PRKCA、JAK1、HSPA1A、PLCB1、GNG4、SHC1和CXCL1基因的表达水平与卵巢癌和肺腺癌患者生存率之间的关系。仅GNG4和PRKCA基因的表达改变与两种肿瘤患者的生存率呈负相关(P<0.05),即GNG4和PRKCA高表达患者的生存率均明显低于低表达患者,见图4。

A:人卵巢上皮癌顺铂耐药细胞株A2780/DDP;B:人肺腺癌顺铂耐药细胞株A549/DDP;a:P<0.05,与A2780比较;b:P<0.01,与A2780比较;c:P<0.05,与A549比较;d:P<0.01,与A549比较

A:GNG4的表达与卵巢癌患者总生存率的Kaplan-Meier曲线;B:PRKCA的表达与卵巢癌患者总生存率的Kaplan-Meier曲线;C:GNG4的表达与肺腺癌患者总生存率的Kaplan-Meier曲线;D:PRKCA的表达与肺腺癌患者总生存率的Kaplan-Meier曲线
3 讨 论
卵巢癌与肺癌是严重威胁人类生命健康的恶性肿瘤。在过去几十年里,尽管分子靶向治疗等新型治疗方案的发展大大提高了患者的生存率,但靶向治疗往往存在适用范围小、治疗费用高等问题,因此以铂类药物为基础的传统化学疗法仍然是大多数卵巢癌及肺癌患者最主要的治疗手段。而肿瘤细胞顺铂耐药性的产生,则是导致化疗失败最主要的原因。现有研究显示,肿瘤细胞对顺铂耐药的形成是多方面的,包括铂的聚集、铂的失活、DNA损伤修复、细胞衰老、凋亡、自噬及上皮间质转化等,几乎涵盖了调节细胞生存的所有方面,且往往是多因素同时存在的,这也是目前缺乏有效逆转肿瘤细胞对铂类药物耐药的主要原因。研究表明,基因的表达改变是肿瘤细胞耐药发生的重要基础,因此,利用基因芯片筛选出与顺铂耐药相关的关键基因,可作为初步阐明顺铂耐药机制的重要手段。本研究通过GEO数据库中卵巢上皮癌及肺癌顺铂耐药与亲本敏感细胞株的基因芯片表达数据集筛选得到295个表达上调和176个表达下调的共同差异基因,提示这些基因可能与顺铂耐药的形成相关。GO与KEGG富集分析结果显示,共同差异基因主要富集于细胞损伤修复、Th17细胞分化、PI3K-Akt信号通路、细胞黏附、轴突导向和细胞因子间受体相互作用等主要与顺铂耐药形成相关的DNA损伤修复和细胞凋亡等过程及通路。
本研究通过在线数据库STRING对共同差异基因进行蛋白互作分析,筛选得到节点较多的10个关键基因,其中9个在耐药株中的表达高于其亲本敏感株,分别为RIPK4、JUN、PRKCA、JAK1、HSPA1A、HSPB1、PLCB1、GNG4、SHC1,而CXCL1在耐药株中的表达则低于敏感株。通过qRT-PCR检测发现除HSPB1外,其余9个关键基因表达情况与芯片结果一致。RIPK4是一个受体相互作用蛋白激酶,研究表明,RIPK4能增强Wnt信号通路从而增强肿瘤细胞的增殖能力并降低肿瘤细胞对顺铂等药物的敏感性,减少顺铂诱导的细胞凋亡进而导致耐药[8]。JUN是AP1复合物的重要组成部分且在多种肿瘤细胞中过表达,它可通过不同机制拮抗p53的促凋亡活性,这也可能是高表达JUN导致肿瘤细胞对顺铂敏感性降低的重要原因[9]。PRKCA是蛋白激酶C家族的成员。FU等[10]研究表明,过表达PRKCA可通过激活粘着斑激酶(FAK)和下游信号如Ras和PI3K/AKT导致细胞对顺铂耐药。JAK1则属于非受体酪氨酸激酶家族,它的底物为STAT,JAK/STAT通路的持续活化可通过上调抗凋亡基因的表达抑制顺铂诱导的肿瘤细胞凋亡,进而导致细胞耐药[11]。HSPA1A属于热休克蛋白家族,研究者发现它们的表达升高与肿瘤患者的不良预后及对顺铂的抗性相关[12]。PLCB1属于PLC酶家族,SENGELAUB等[13]研究发现,PLCB1高表达可促进肿瘤细胞的迁移。而GNG4被认为是一个潜在的肿瘤治疗靶点,PAL等[14]研究认为其高表达同样可促进肿瘤细胞的迁移。有关肿瘤细胞迁移与顺铂耐药形成之间的关系尚未明确,但WACLAW等[15]研究指出,细胞的迁移性可能是影响肿瘤耐药性的关键因素。SHC1在多种肿瘤细胞中发挥致癌作用。AN等[16]研究表明,SHC1的过表达可显著抑制凋亡,导致肿瘤细胞的持续增殖和耐药。以上结果初步表明,在耐药株中高表达的关键基因都可能导致顺铂耐药的产生。而低表达的CXCL1是一个趋化因子,ACHARYYA等[17]发现,阻断CXCL1的表达能提高化疗的有效性,而芯片结果却显示,CXCL1在敏感株细胞中的表达明显高于耐药株,提示CXCL1在卵巢上皮癌及肺腺癌顺铂耐药的形成中可能不起主要作用,这也充分说明了顺铂耐药的形成是一个由多因素、多因子参与的复杂过程。
以上结果表明,在卵巢上皮癌及肺腺癌耐药细胞株中RIPK4、JUN、PRKCA、JAK1、HSPA1A、PLCB1、GNG4、SHC1基因的高表达可能是其耐药形成的重要原因。为了探究耐药相关基因的表达是否影响卵巢癌及肺腺癌患者的生存率,本研究对上述8个耐药相关基因进行了生存分析,结果显示,仅PRKCA和GNG4基因的表达与卵巢上皮癌及肺腺癌患者的总生存率呈负相关,表明当靶向PRKCA和GNG4使其表达下调时,不但能提高卵巢上皮癌及肺腺癌对顺铂的敏感性,还能提高患者的生存率,改善患者的预后。而当靶向其他高表达的关键基因时,虽具有逆转顺铂耐药的潜能,但并不能改善患者的预后。
综上所述,本研究应用生物信息学方法综合分析并验证了卵巢上皮癌与肺腺癌顺铂耐药的共同差异基因,结果得到了8个关键基因RIPK4、JUN、PRKCA、JAK1、HSPA1A、PLCB1、GNG4、SHC1,提示这8个基因可能在多种以顺铂为一线化疗药物的肿瘤耐药形成中起关键作用,为从分子水平更深入探究顺铂耐药发生、发展的共同机制提供了重要思路。此外,GNG4和PRKCA基因的表达还与提高卵巢癌及肺腺癌患者总生存率密切相关,提示其可能作为逆转顺铂耐药和改善患者预后的生物学新靶标。
