土荆芥化感胁迫下蚕豆叶绿体超微结构和光合关键基因表达的变化
2019-01-18阿的鲁骥马丹炜
阿的鲁骥, 周 健,李 洁,马丹炜
(1. 四川师范大学生命科学学院,四川 成都 610101;2. 西南民族大学青藏高原研究所,四川 成都 610064;3.四川省农业科学院农业信息与农村经济研究所, 四川 成都 610066)
【研究意义】植物的光合作用是一系列复杂代谢反应的总和。由于步骤繁多、过程复杂,任何一个反应步骤受到影响都有可能影响到光合作用。生物体的所有生长发育过程都受基因表达的调控,同样光合作用也是相关基因表达调控的结果[1]。【前人研究进展】光合作用过程对环境变化十分敏感,在逆境胁迫下,植物光合产物减少,植物生长发育受阻[2]。生物入侵已经成为导致全球生物多样性下降的一个重要因素,扩散到新生境的入侵植物比原产地的个体具有更强的化感作用,使其在与其他种群争夺环境资源的竞争中获得优势地位[3]。入侵植物释放到周围的化感物质具有较大的细胞毒性[4],这些物质通过细胞膜进入细胞后往往会破坏细胞的内部结构[5],并干扰受体植物的基因表达[6]。入侵植物土荆芥(ChenopodiumambrosioidesL.)释放的挥发油及其主要成分α-萜品烯和对伞花素诱导周围植物的根细胞[7]和气孔保卫细胞[8]凋亡,并干扰了根尖细胞的抗氧化酶活性及其基因表达[9]。【本研究切入点】有关土荆芥挥发性化感物质是否影响与光合作用密切相关的细胞器叶绿体的结构以及光合关键基因表达的研究却鲜有报道。【拟解决的关键问题】本研究采用盆栽试验,以土荆芥入侵地广泛栽培的农作物蚕豆(ViciafabaL.)为受体,运用透射电镜技术和RT-PCR技术,研究了土荆芥挥发油及其主要成分α-萜品烯和对伞花素对受体植物幼苗叶片叶绿体超微结构和光合相关基因表达的影响,以期为深入探讨土荆芥的入侵机制提供理论依据。
1 材料与方法
1.1 材料
供试土荆芥挥发油由本实验室提供。色淡黄,密度843 mg/mL,其主要成分α-萜品烯和对伞花素的含量分别为151和156 mg/mL;α-萜品烯和对伞花素的标准品购自成都市锐可思生化试剂公司;蚕豆种子(成胡14#)购于成都市五块石种子市场。
1.2 幼苗培养和化感胁迫处理
幼苗培养方法见本项目组前期研究[4]。待蚕豆幼苗长至4~5周进行化感胁迫处理;本研究共设置3个处理组(挥发油处理组、α-萜品烯处理组、对伞花素处理组)和1个对照组。在预实验基础上确定挥发油的处理剂量,将该处理剂量挥发油中对伞花素和α-萜品烯的含量定为二者的处理剂量;挑选长势一致、生长良好的蚕豆幼苗若干盆,随机分为4组,分别置于特制的、带有密封盖子的玻璃箱(长25 cm、宽20 cm、高40 cm)中,每箱均匀放置5盆,在每个箱底部正中央放置一个小培养皿;分别将80 μl挥发油、14.88 μl α-萜品烯和3.52 μl对伞花素滴加在玻璃箱中央的小培养皿中,迅速盖上盖子,并用凡士林封住玻璃箱口。对照组不作处理但同样用凡士林封住玻璃箱口。将4组玻璃箱均置于25 ℃光照培养箱中处理3 h取出挥去挥发物,再放入培养箱中培养。每天处理时段为10:00-13:00,连续处理3 d,第4天取样电镜观察和real-time PCR分析。
1.3 叶绿体超微结构的观察
取处理第4天后的叶片,在中脉两侧剪取1 mm×1 mm的小块若干,2.5 %戊二醛前固定,0.1 mol/L PBS(pH 7.4)漂洗,1 %锇酸后固定, 0.1 mol/L PBS(pH 7.4)漂洗,乙醇梯度系列脱水,丙酮︰Epon 812包埋剂=1∶1的混合液渗透,Epon-812环氧树脂包埋,Leica UC7切片机切片,铀铅双染色(2 %醋酸铀饱和水溶液和柠檬酸铅分别染色15min),室温干燥过夜。Tecnai G2 20 TWIN透射电子显微镜观察并采集图像。
1.4 叶片RNA的提取与RT-PCR分析
总RNA的提取: 取处理第4天后的叶片,用液氮充分研磨,按照天根生化科技有限公司的植物总RNA提取试剂盒操作说明提取蚕豆幼苗叶片的总RNA,除去DNA后分装于EP管中,-80 ℃保存待测。
引物设计:用Primer Premier 5.0引物设计软件,设计蚕豆核酮糖羧化酶基因(Rbcs-3C)、光系统ⅡCP47蛋白基因(PsbB)、ATP合成酶β亚基基因(beta)、质体蓝素基因(PetE)和内参基因(EF-1-alpha)特异性引物,序列如下。EF-1-alphaF:5′-ACGAGGCTCTCACTGAGGCTCTTCC-3′;EF-1-alphaR:5′-CCTTGGCAGGGTCATCCTTGGAGTTG-3′;Rbcs-3CF:5′-CCGTGAGCACAACAAGTCACCAGGAT-3′;Rbcs-3CR:5′-GGTAAGCCGCAACAACTTCATCAAGC-3′;PsbBF:5′-CCGCTCTAACAATTCCGTCTCCGTCT-3′;PsbBR:5′-CGAAGAGTTGGTGCTGGGCTAGTCAA-3′;betaF:5′-TTCTTCACCGACGATGCGAGGTTGGA-3′;betaR:5′-GACTGATCCTGCTCCTGCCACGACAT-3′;PetEF:5′-GCTGTAAGTCTCACCAGGAGCGTTGA-3′;PetER:5′-AGCAATGCCTTGGCTGTTGAAGTCTT-3′。
引物均由上海生工生物工程技术服务有限公司合成。
RT-PCR分析:使用实时荧光定量PCR仪(PIKORed 96,美国Thermo Fisher)进行RT-PCR分析。反应体系为20 μl,含SYBR Primix ExTaqⅡ10 μl, PCR Forward Primer 0.8 μl,PCR Reverse Primer 0.8 μl,cDNA 2 μl和ddH2O 6.4 μl;荧光 PCR 反应程序为:95 ℃ 1 min,95 ℃ 5 s,62 ℃ 30 s,45个循环;每处理重复3次;使用Thermo Scientific PikoReal软件分析PCR过程的CT(Threshold cycle)值。
1.5 数据统计
参照Williamson et al.[10]的方法计算基因相对表达量的化感敏感指数(RI):RI=1-C/T(T≥C);RI=T/C-1(T 用透射电子显微镜观察发现,对照组中的蚕豆叶片叶绿体的形态结构正常(图1 a, e, i),叶绿体长梭形,膜结构清晰完整,排列有序,并紧贴于细胞壁,叶绿体中基质均匀且浓密,基粒类囊体垛叠整齐紧密,淀粉粒发育正常;挥发油处理组(图1d, h, l)中,叶绿体脱离细胞壁散乱分布于细胞中央,结构紊乱,表现为肿胀、膜结构受损明显,叶绿体内部基质不均匀、基粒类囊体排列紊乱、淀粉粒消失、片层扭曲且排列松散;α-萜品烯处理组中(图1 c, g, k),叶绿体出现肿胀现象,部分叶绿体脱离细胞壁,叶绿体膜和内部结构变化不明显;对伞花素处理组(图1 b, f, j)中,叶绿体超微结构与对照组相比无差异,表明对伞花素对蚕豆叶绿体超微结构没有产生影响。 a, e, i: 对照组;b, f, j: 对伞花素处理组;c, g, k: α-萜品烯处理组;d, h, l: 挥发油处理组;CH:叶绿体;SG:淀粉粒;CW:细胞壁;Me:叶绿体膜;G:基粒;GL:基粒片层;N:细胞核a, e and i are control groups;b, f and j were cymene-treated groups; c, g and k are α-terpinene-treated groups;CH is Chloroplast; SG is Starch Grain; CW is Cell Wall; ME is Chloroplast Membrane; G is Grana; GL is Grana Lamella; N is Nucleus图1 土荆芥挥发油、α-萜品烯和对伞花素作用下蚕豆幼苗叶片超微结构Fig.1 TEM micrographs of leave of Vicia faba exposure to the volatile oil from Chenopodium ambrosioides, α-terpinene and cymene 不同小写字母表示P<0.05 的显著水平Different lowercase letters indicates P<0.05图2 土荆芥挥发油、α-萜品烯和对伞花素对蚕豆幼苗Rbcs-3C、PsbB、beta和PetE 基因相对表达量的影响Fig.2 Effects of volatile oil from Chenopodium ambrosioides L., α-terpinene and cymene on relative expression of Rbcs-3C, PsbB, beta and PetE in Vicia faba L. RT-PCR研究表明,在土荆芥挥发油、α-萜品烯和对伞花素作用下,蚕豆幼苗光合关键基因的表达发生了不同程度的变化(图2)。其中,挥发油处理组中蚕豆幼苗光合关键基因表达的变化幅度最大,核酮糖羧化酶基因(Rbcs-3C)、光系统ⅡCP47蛋白基因(PsbB)、ATP合成酶β亚基基因(beta)、质体蓝素基因(PetE)等4个基因的相对表达量均显著高于对照组(P<0.05);与对照组相比,对伞花素处理组的Rbcs-3C和PsbB的相对表达量均下调,beta和PetE的相对表达量均上调,但只有PetE相对表达量与对照组的差异达到显著水平(P<0.05);α-萜品烯处理组中,PsbB的相对表达量低于对照组,但未达到显著水平(P>0.05),而Rbcs-3C、beta和PetE的相对表达量均显著高于对照组(P<0.05)。由此表明,土荆芥挥发性化感物质不同程度影响了受体植物光合关键基因的表达,无疑会干扰受体植物的光合作用过程。 从图2-A可见,挥发油处理组和α-萜品烯处理组之间Rbcs-3C的表达量差异不显著(P>0.05),但二者的相对表达量均显著高于对伞花素处理组(P<0.05);从图2-B可知,PsbB的相对表达量在对伞花素处理组和α-萜品烯处理组之间无显著差异(P>0.05),但二者与挥发油处理组之间的差异达到显著水平(P<0.05);3个处理组的beta相对表达量均呈现显著差异(P<0.05,图2-C),而PetE的相对表达量则无显著差异(P>0.05,图2-D)。 3种挥发物对蚕豆幼苗光合关键基因化感作用的综合效应分别为对伞花素0.525,α-萜品烯0.535,挥发油0.666,由此可见,挥发油对光合关键基因的相对表达量影响最大,而挥发油主要成分α-萜品烯和对伞花素的化感作用强度不及挥发油,说明挥发油中多种成分存在正向的协同效应。 4种基因相对表达量对土荆芥挥发油的敏感程度从大到小依次为PetE,Rbcs-3,CPsbB和beta。 叶绿体是植物体内叶绿素存在的部位,是进行光合作用的场所[11]。化感作用对叶绿体等细胞器超微结构的影响将直接改变受体植物的生长发育。叶绿体结构受损时,叶绿素合成酶活性受抑制,叶绿素降解酶活性增强,导致叶绿素的含量下降[12]。项目组前期研究发现,在土荆芥挥发油及其两个主要成分α-萜品烯和对伞花素作用下,蚕豆幼苗的最大光化学效率(Fv/Fm)、实际光化学效率(ΦPsⅡ) 和叶绿素含量均呈现下降趋势,土荆芥化感胁迫可能干扰了叶绿素的合成过程或引起叶绿素降解,从而降低了光合速率[13]。本研究结果表明,在土荆芥挥发油、α-萜品烯和对伞花素的作用下,蚕豆幼苗叶片的叶绿体超微结构受到不同程度的破坏,其中挥发油处理组中叶绿体受损程度最为严重,叶绿体散乱分布于细胞中央,结构紊乱,表现为肿胀、膜结构受损明显,叶绿体内部基质不均匀、基粒类囊体排列紊乱、淀粉粒消失、片层扭曲且排列松散。相对而言,对伞花素对叶绿体的损害较小。由此可见,土荆芥化感胁迫抑制邻近植物的生长发育[14-15],无疑与其化感物质破坏了叶绿体超微结构、影响光合作用过程有关。 化感物质通过上调或下调某些基因,或诱导新基因的表达,影响受体植物相关基因的表达调控[16]。例如,珍珠绣线菊(SpiraeathunbergiiBl.)释放的顺式肉桂酸改变了拟南芥(ArabidopsisthalianaL.)237个基因的表达量[17],土荆芥挥发性化感物质下调了玉米(ZeamaysL.)抗氧化酶基因的表达[9]。光合作用涉及多个相互协调且复杂的反应步骤,其每一步反应均容易受到各种环境胁迫的影响[18-19]。本研究结果表明,在挥发油、α-萜品烯和对伞花素作用下,与光合作用有关的关键基因表达发生了不同程度的变化。整体来看,大部分处理中,基因表达表现为上调,仅仅3个处理的基因表达下调(对伞花素处理组的Rbcs-3C和PsbB;α-萜品烯处理组的PsbB)。PsbB和PetE的表达产物是光合电子传递链的重要成员,beta的表达产物与叶绿体ATP的合成有关,而Rbcs-3C表达异常将直接改变受体植物的碳同化过程。本研究结果显示,土荆芥释放到环境中的挥发性化感物质干扰了蚕豆幼苗光合关键基因的正常表达,打破了蚕豆幼苗的光合作用过程各个环节协调性,进而影响了幼苗的正常生长。 土荆芥挥发油的成分十分复杂[22],这些成分在植物化感作用中的地位不尽相同,且可能相互之间可能存在着拮抗效应或协同效应。从本研究结果可见,挥发油对叶绿体结构的损害和对光合作用关键基因表达的影响最大,α-萜品烯次之,对伞花素的影响最小。这一结果与本项目组前期的研究结果相同[7-9, 20-21]。综合分析可知,土荆芥向周围环境释放的挥发性化感物质之间存在着协同作用,进而使挥发油的化感效应大于其主要成分α-萜品烯和对伞花素。 在土荆芥挥发油、α-萜品烯、对伞花素作用下,蚕豆幼苗叶片叶绿体的超微结构受损,叶绿体脱离细胞壁散乱分布于细胞中央,结构紊乱,表现为肿胀、膜结构受损明显,叶绿体内部基质不均匀、基粒类囊体排列紊乱、淀粉粒消失、片层扭曲且排列松散;核酮糖羧化酶基因(Rbcs-3C)、光系统ⅡCP47蛋白基因(PsbB)、ATP合成酶β亚基基因(beta)、质体蓝素基因(PetE)等光合关键基因的表达异常。综上所述,破坏受体植物叶绿体结构、干扰光合作用关键基因表达是土荆芥化感胁迫的机制之一。2 结果与分析
2.1 土荆芥挥发油、α-萜品烯和对伞花素对蚕豆幼苗叶绿体超微结构的影响

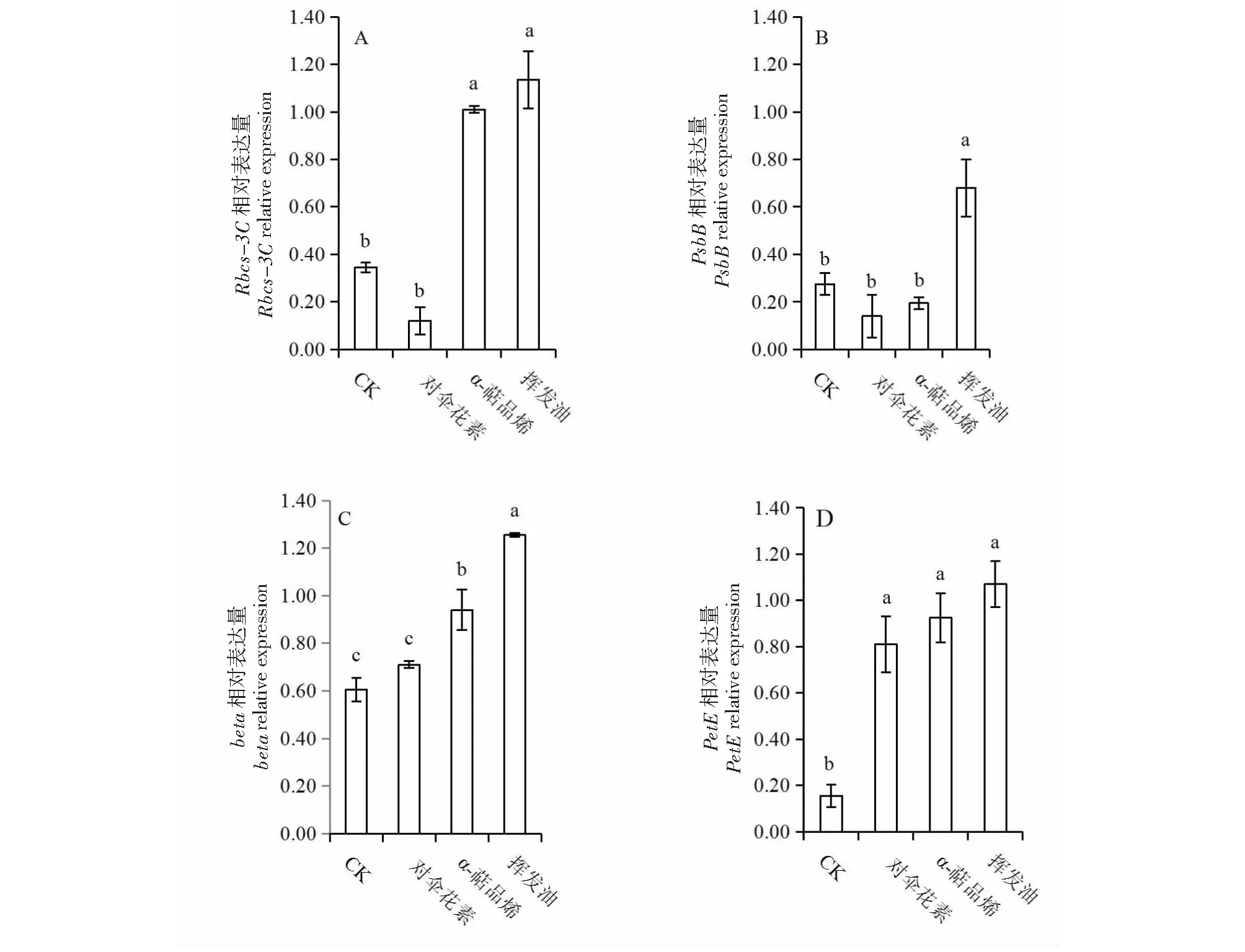
2.2 土荆芥挥发油、α-萜品烯和对伞花素对蚕豆幼苗光合作用相关基因表达量的影响
3 讨 论
4 结 论
