丛枝菌根真菌对镍胁迫桂花幼苗光合作用及抗氧化酶活性的影响
2019-01-03李晓曼王建军
李晓曼 王建军
摘要:以桂花幼苗为材料,研究接种丛枝菌根真菌(AMF)对0、100、200、300、400 mg/L这5种镍(Ni)质量浓度胁迫下桂花光合作用及抗氧化酶活性等的影响。结果表明,5种Ni胁迫质量浓度下,未接种AMF的桂花幼苗根系菌根侵染率均为0,而接种AMF的根系菌根浸染率则随Ni胁迫质量浓度的增大而逐渐降低;随Ni胁迫质量浓度的增大,接种、未接种AMF的桂花幼苗叶片叶绿素含量、类胡萝卜素含量、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、植株株高、茎粗、总干物质含量呈逐渐降低趋势,且接种AMF的桂花幼苗相应指标下降幅度多显著低于未接种处理(P<0.05),而胞间二氧化碳浓度(Ci)呈先降后增,接种AMF的桂花幼苗叶片Ci显著低于未接种处理,相对电导率、丙二醛(MDA)含量呈逐渐上升趋势,且接种AMF的桂花幼苗的相应指标多显著低于未接种处理,超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)则呈先升后降趋势,接种AMF的桂花幼苗抗氧化酶活性显著高于未接种处理;Ni胁迫质量浓度为400 mg/L时,接种AMF的桂花幼苗叶绿素含量、Pn、Gs、Tr、株高、总干物质质量、SOD活性、POD活性、CAT活性分别较未接种的分别显著增加69.72%、80.22%、61.90%、42.48%、22.12%、15.76%、6085%、21.83%、22.03%,Ci、相对电导率、MDA含量分别较未接种的显著降低6.98%、10.04%、19.21%。
关键词:丛枝菌根真菌;桂花;光合作用;抗氧化酶;镍
中图分类号: S685.130.1;S182文献标志码: A
文章编号:1002-1302(2019)21-0223-05
收稿日期:2018-08-07
基金项目:全国农业职业教育“十三五”科研项目(编号:2016-135-Y-005)。
作者简介:李晓曼(1980—),女,辽宁营口人,硕士,讲师,从事园林植物栽培及园林规划设计研究。E-mail:582659554@qq.com。
近年来,随着我国工业化的飞速发展,土壤环境污染问题日趋严重,其中,土壤重金属污染因其毒性强、易富集、难降解等特点而愈发被人们关注[1-2]。据《全国土壤污染状况调查公报》显示,我国土壤重金属总超标率为16.1%,其中,镍(Ni)超标量达到4.8%,仅次于镉(Cd),已成为我国土壤重金属污染的主要元素之一[3]。镍是植物生长发育所必需的微量元素之一,但当土壤中Ni含量过高时,往往会对植物水分、养分的吸收及光合作用产生干扰,进而影响农作物产量、品质及园林绿化植物的景观效果,而如何提高植物对Ni胁迫的耐受性已成为亟待解决的关键问题之一[4-5]。
丛枝菌根真菌(arbuscular mycorrhiza fungi,AMF)是一类在土壤中广泛存在的有益真菌,可与绝大多数植物形成互利的共生体系,在提高植物对生物及非生物胁迫抗性方面发挥着非常重要的作用[6-7]。有研究表明,外源接种AMF可提高植物对重金属的耐受性,其作用机制主要体现在:一是AMF可通过菌丝体组分纤维素、几丁质等来吸附重金属,减少重金属在植物体内的转移量[8-9];二是AMF可对植物的根系分泌物、根际土壤微生物群落结构及土壤pH值产生影响,从而降低土壤重金属的有效性,减少植物吸收[10-11];三是AMF可以改善植物礦质营养和根系形态,从而影响植物对重金属的吸收和转移[12-13];四是AMF可以提高植物的抗氧化酶活性,从而提高植物对重金属的耐受性[14-15]。目前,有关提高植物对镍耐受性的方法主要为化学方法,如外源施用甜菜碱、褪黑素、水杨酸等[3,16-17],但存在成本高、周期长且易造成二次污染等缺点。
桂花(Osmanthus sp.)别称岩桂,为木犀科木犀属植物,是我国十大名花之一,因其集绿化、香化、美化于一体,在我国园林绿化中被广泛应用,而当前利用AMF提高其对镍耐受性的研究鲜见报道。本研究以桂花幼苗为试验材料,采用不同质量浓度Ni进行胁迫处理,并分别接种AMF,以分析接种AMF对镍胁迫下桂花幼苗光合作用及抗氧化酶活性的影响,以期为提高植物对镍胁迫的耐受性提供一定的理论依据。
1材料与方法
1.1试验材料
试验于2017年4月在重庆三峡职业学院苗圃进行,供试植物为桂花幼苗,购自重庆科光种苗有限公司;供试丛枝菌根真菌为摩西球囊霉菌(Glomus mosseae),购自北京市农林科学院植物保护环境保护研究所;氯化镍(NiCl2)分析纯,购自上海阿拉丁生化科技股份有限公司。
1.2试验方法
桂花幼苗采用直径为40 cm、深45 cm的花盆栽种,每盆定植1株,栽培基质为改良土壤基质,定植前,以基质接种丛枝菌根真菌20 g为处理(AMF),以接种经高温灭活的丛枝菌根真菌20 g(视为未接种丛枝菌根真菌,全文同)为对照(CK);缓苗生长30 d,即2017年6月,选取长势、大小基本一致的桂花幼苗,分别采用NiCl2进行Ni胁迫试验,设NiCl2终质量浓度分别为0、100、200、300、400 mg/L,每天按100 mg/L梯度递增,直至达到各处理预定质量浓度;后每隔3 d按预定镍质量浓度浇灌1次,浇灌量为250 mL;连续镍胁迫处理60 d,采样测定各指标。试验共设10个处理,每处理20盆,随机区组排列,试验期间进行正常养分管理及病虫害防治。
1.3测定内容与方法
1.3.1桂花幼苗生长及菌根侵染情况测定胁迫处理结束,采用软尺测量桂花植株株高、茎粗,采用电子天平称量总的干物质质量;桂花幼苗根系菌根侵染情况采用Phillips等的方法[18]测定,统计菌根侵染率,计算公式为:
菌根侵染率=侵染菌根段数/检测菌根段数×100%。
1.3.2光合指标及抗氧化酶活性测定每处理选取植株的第2~3张功能叶,采用美国LI-COR公司产Li-6400 XT型便携式光合测量仪测定其净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr);采用丙酮比色法测定光合色素含量,采用相对电导法(RC)测定叶片细胞膜透性;分别采用抑制氮蓝四唑(NBT)光还原比色法、愈疮木酚法和紫外分光光度法测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性;采用硫代巴比妥酸比色法测定丙二醛(MDA)含量。
1.4数据分析
采用Excel 2013软件对数据进行整理、统计及作图,采用SPSS 18.0软件进行差异显著性分析。
2结果与分析
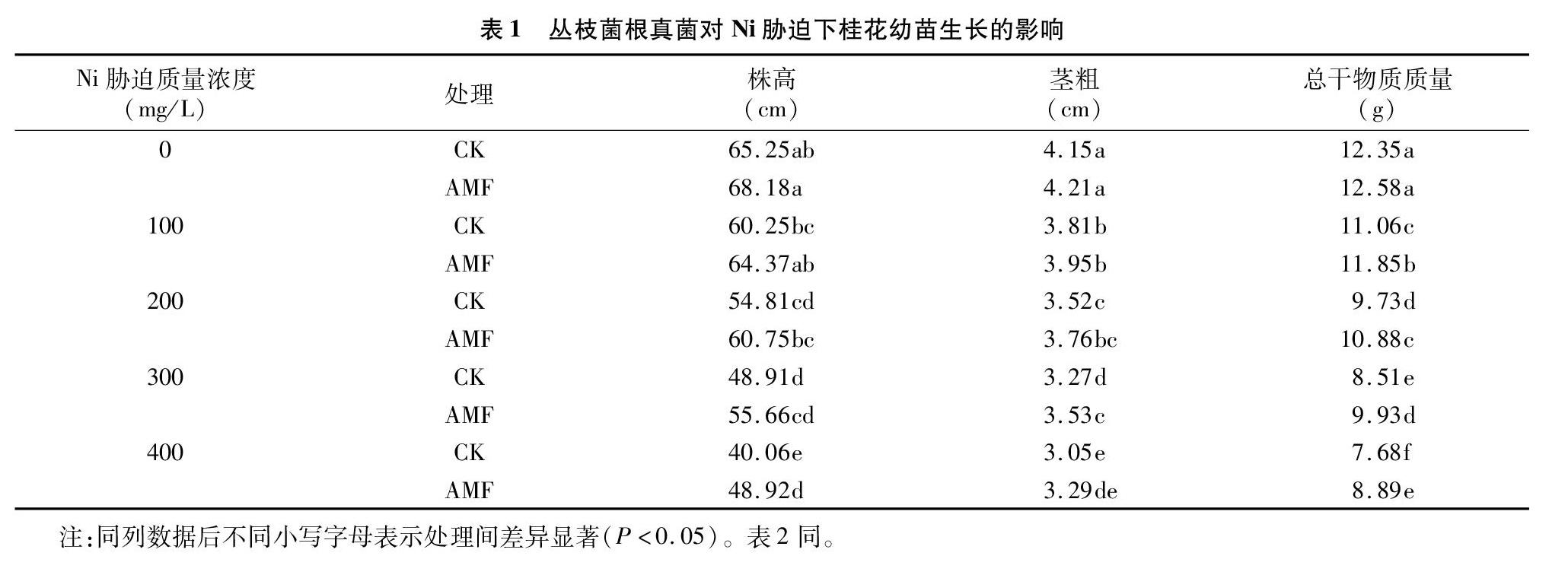
2.1丛枝菌根真菌对Ni胁迫下桂花幼苗生长的影响
由表1可见,随着Ni胁迫质量浓度的增大,接种、未接种AMF的桂花幼苗植株株高、茎粗、总干物质质量均呈逐渐减小趋势,接种AMF的植株株高、茎粗、总干物质质量均明显高于未接种处理;Ni胁迫质量浓度分别为100、200、300、400 mg/L 时,接种AMF的桂花幼苗株高较CK处理分别增加6.84%、10.84%、13.80%、22.12%,除Ni胁迫质量浓度为400 mg/L 的植株株高显著高于未接种的(P<0.05)外,其他3种质量浓度处理接种与未接种间差异不显著(P>0.05),接种AMF的桂花幼苗茎粗分别较CK处理增加3.67%、682%、7.95%、7.87%,除Ni胁迫质量质量浓度为300 mg/L的植株幼苗茎粗显著高于未接种的外,其他3种质量浓度处理接种与未接种间差异不显著,接种AMF的桂花幼苗总干物质质量分别较CK处理增加7.14%、11.82%、16.69%、1576%,接种与未接种间差异显著。可见,Ni胁迫可明显抑制桂花幼苗的生长,而接种AMF可明显促进Ni胁迫下桂花幼苗的生长。
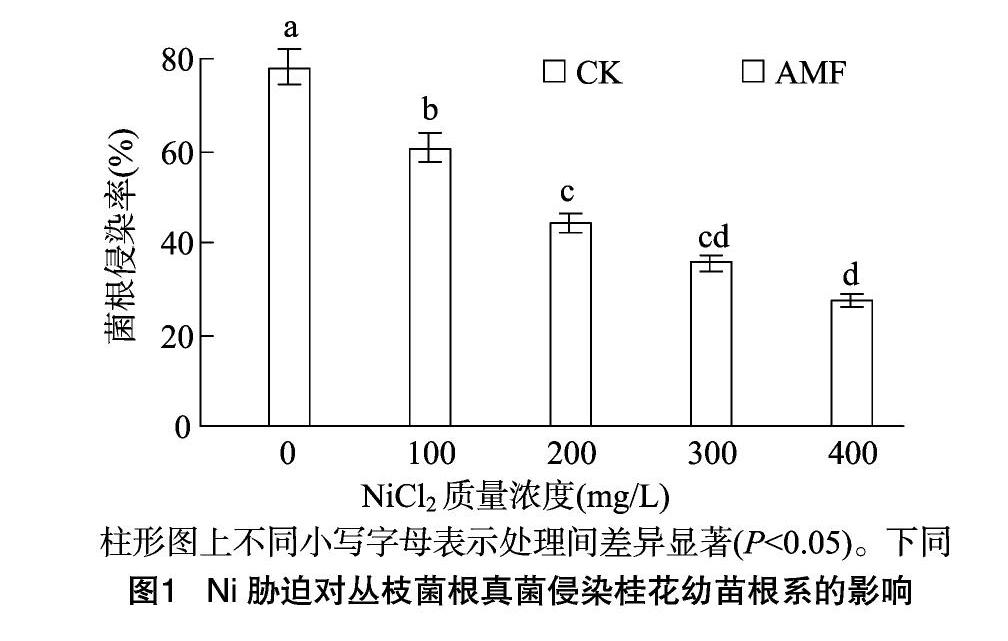
2.2Ni胁迫对丛枝菌根真菌侵染桂花幼苗根系的影响
由图1可见,5种Ni质量浓度胁迫下,未接种丛枝菌根真菌的桂花幼苗根系均未发现有菌丝生成,菌根侵染率为0;随Ni胁迫质量浓度的增大,接种AMF的桂花幼苗根系菌根浸染率逐渐降低,可能是由于Ni胁迫不利于AMF的侵染和菌根发育所致;Ni胁迫质量浓度为0 mg/L时,接种AMF的桂花幼苗根系菌根浸染率为78.25%,而Ni胁迫质量浓度分别为100、200、300、400 mg/L时,接种AMF的桂花幼苗根系菌根浸染率分别为60.58%、44.39%、35.68%、27.22%,分别较0 mg/L处理的显著降低17.67、33.86、42.57、51.03百分点(P<0.05)。
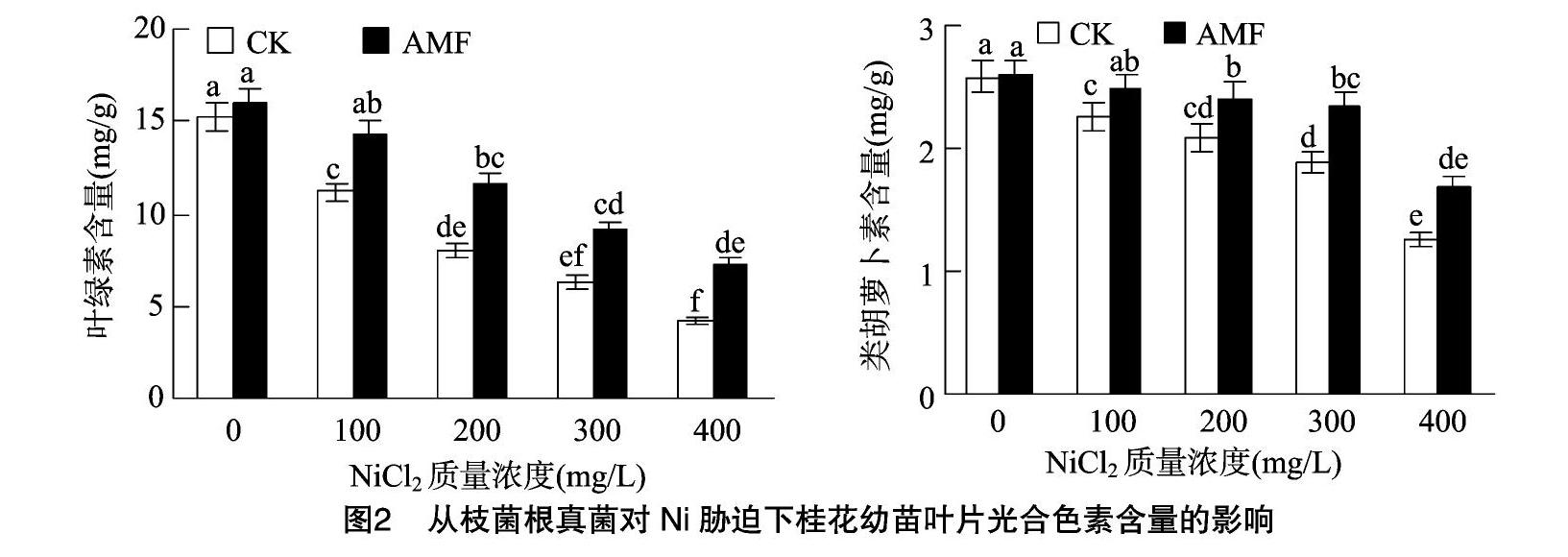
2.3丛枝菌根真菌对Ni胁迫下桂花幼苗叶片光合色素含量的影响
由图2可见,随Ni胁迫质量浓度的增大,接种、未接种AMF的桂花幼苗叶片叶绿素、类胡萝卜素含量均呈逐渐下降趋势 接种AMF的叶绿素、类胡萝卜素含量均明显高于未接
种处理;Ni胁迫质量浓度分别为100、200、300、400 mg/L时,接种AMF的桂花幼苗叶片叶绿素含量分别为14.25、11.69、9.05、7.23 mg/g,分别较未接种的显著增加27.80%、4522%、44.34%、69.72%(P<0.05);Ni胁迫质量浓度分别为100、200、300 mg/L时,接种AMF的桂花幼苗叶片类胡萝卜素含量分别为2.48、2.41、2.33 mg/g,分别较未接种的显著增加10.22%、15.31%、23.94%,而Ni胁迫质量浓度为400 mg/L 时,接种AMF的桂花幼苗叶片类胡萝卜素含量为1.68 mg/g,较未接种的增加33.33%,但相互间差异不显著(P>0.05)。可見,Ni胁迫可明显降低桂花幼苗叶片光合色素含量,而接种AMF可明显提升Ni胁迫下桂花幼苗叶片的光合色素含量。
2.4丛枝菌根真菌对Ni胁迫下桂花幼苗叶片光合作用的影响
由图3可知,随Ni胁迫质量浓度的增大,接种、未接种AMF的桂花幼苗叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)均呈逐渐下降趋势,且接种AMF的Pn、Gs、Tr均明显高于未接种处理,而接种、未接种AMF的桂花幼苗叶片胞间二氧化碳浓度(Ci)呈先降后升趋势,接种AMF的Ci显著低于未接种处理;Ni胁迫质量浓度分别为100、200、300、400 mg/L 时,接种AMF的桂花幼苗叶片Pn分别为15.55、13.15、9.98、6.65 μmol/(m2·s),分别较未接种的增加1013%、18.58%、32.01%、80.22%,除Ni胁迫质量浓度为300 mg/L的叶片Pn与未接种的差异不显著(P>0.05)外,其他3种质量浓度处理下接种AMF的叶片Pn显著高于未接种的(P<0.05),接种AMF的桂花幼苗叶片Gs分别为241、201、153、102 mmol/(m2·s),分别较未接种的显著增加711%、10.44%、33.04%、61.90%(P<0.05),接种AMF的桂花幼苗叶片Tr分别为3.12、2.71、2.35、2.18 mmol/(m2·s),分别较未接种的显著增加11.03%、15.32%、19.90%、4248%;Ni胁迫质量浓度分别为100、400 mg/L时,接种AMF的桂花幼苗叶片Ci分别为260.35、358.69 μmol/mol,分别较未接种的显著降低8.88%、6.98%。可见,Ni胁迫可明显降低桂花幼苗叶片的Pn、Gs、Tr,增加Ci,光合作用受到明显抑制,而接种AMF可明显缓解Ni胁迫对桂花幼苗叶片光合作用的抑制。
2.5丛枝菌根真菌对Ni胁迫下桂花幼苗叶片抗氧化酶活性的影响
由表2可见,随Ni胁迫浓度的增大,接种、未接种AMF的桂花幼苗叶片SOD、 POD、CAT活性均呈先升后降趋势,除Ni胁迫质量浓度为0 mg/L外,接种AMF的SOD、POD、CAT活性均显著高于未接种处理(P<0.05);Ni胁迫质量浓度为300 mg/L时,接种AMF的桂花幼苗叶片SOD、POD、CAT活性分别较未接种的显著增加29.58%、32.59%、24.93%,Ni胁迫质量浓度为400 mg/L时,接种AMF的桂花幼苗叶片SOD、POD、CAT活性分别较未接种的显著增加60.85%、2183%、22.03%。可见,接种AMF可明显提升Ni胁迫下桂花幼苗叶片的抗氧化酶活性,提升抗氧化能力。
2.6丛枝菌根真菌对Ni胁迫下桂花幼苗叶片细胞膜透性的影响
由图4可见,随Ni胁迫浓度的增大,接种、未接种AMF的桂花幼苗叶片相对电导率呈逐渐增加趋势,接种AMF的相对电导率多显著低于未接种处理(P<0.05);Ni胁迫质量浓度分别为100、200、300、400 mg/L时,接种AMF的桂花幼苗叶片相对电导率分别为11.22%、16.75%、24.26%、4065%,分别较未接种的降低2.94、5.26、9.30、10.04百分点,除Ni胁迫质量浓度为100 mg/L的叶片相对电导率与未接种的差异不显著(P>0.05)外,其他3种质量浓度处理下接种AMF的叶片相对电导率显著高于未接种的;随Ni胁迫质量浓度的增大,接种、未接种AMF的桂花幼苗叶片MDA含量呈先降后增趋势;Ni胁迫质量浓度分别为100、200 mg/L时,接种AMF的桂花幼苗叶片MDA含量分别为16.85、1856 nmol/g,分别较未接种的降低0.94%、12.70%,与未接种的差异不显著(P>0.05),而Ni胁迫质量浓度分别为300、400 mg/L时,接种AMF的桂花幼苗叶片MDA含量分别为23.25、31.92 nmol/g,分别较未接种的显著降低18.68%、1921%。这说明,Ni胁迫可对桂花幼苗葉片的细胞膜造成破坏,而接种AMF可显著降低Ni胁迫下桂花幼苗叶片的细胞膜破坏程度,可能是由于接种AMF可明显提升Ni胁迫下桂花幼苗叶片的抗氧化水平,进而降低活性氧水平所致。
3结论与讨论
菌根侵染率是衡量植物根系菌根化的重要指标,侵染率越高说明菌根效应越强[19]。本研究结果表明,随镍(Ni)胁迫质量浓度的增大,接种丛枝菌根真菌(AMF)的桂花幼苗根系菌根侵染率逐渐降低,Ni胁迫质量浓度分别为0、400 mg/L时,接种AMF的桂花幼苗根系菌根侵染率分别为78.25%、27.22%,差异显著(P<0.05),可能是由于重金属胁迫会对丛枝菌根真菌合成和侵染产生一定的抑制作用,这与尹大川等的研究结果[14,19]较为一致。
叶片光合色素是植物进行光合作用的重要色素,其含量高低与植物光合作用强弱密切相关。本研究结果表明,随着Ni胁迫质量浓度的增大,桂花叶片叶绿素、类胡萝卜素含量呈逐渐下降趋势,而接种AMF的桂花其叶绿素、类胡萝卜素含量均明显高于未接种处理。净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间二氧化碳(Ci)是衡量光合作用的重要参数。本研究结果表明,随着Ni胁迫质量浓度的增大,桂花叶片Pn、Gs、Tr呈逐渐降低趋势,而Ci则呈先降后增趋势,接种AMF的桂花幼苗叶片Pn、Gs、Tr明显高于未接种处理、Ci则明显低于未接种处理;Ni胁迫质量浓度为400 mg/L时,接种AMF的桂花幼苗叶片叶绿素含量、Pn、Gs、Tr分别较未接种显著增加69.72%、80.22%、61.90%、42.48%(P<005),而Ci则较未接种显著降低6.98%,这种造成光合速率下降的原因可能是由于Ni胁迫可使叶肉细胞光合活性下降所致,而接种AMF可明显缓解桂花因Ni胁迫而造成的光合色素降解,进而提高光合速率,同时,接种AMF可明显提升Ni胁迫下桂花的叶肉细胞光合活性,从而提高光合速率,这与尹大川等的研究结果[14,19]较为一致。
植物在遭遇生物和非生物胁迫时,会启动自身的抗氧化防御体系,以最大程度地减少逆境对自身造成的伤害[20]。本研究结果表明,随Ni胁迫质量浓度的增大,桂花叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性均呈先增后降趋势,而接种AMF的SOD、POD、CAT活性均明显高于未接种处理。相对电导率和丙二醛(MDA)含量是衡量膜质过氧化程度的重要指标。本研究结果表明,随Ni胁迫质量浓度的增大,桂花叶片的相对电导率和MDA含量呈增加趋势,且接种AMF的相对电导率、MDA含量均明显低于未接种处理;Ni胁迫质量浓度为400 mg/L时,接种AMF的桂花幼苗叶片SOD、POD、CAT活性分别较未接种的显著增加60.85%、21.83%、22.03%,而相对电导率、MDA含量分别较未接种的显著降低10.04%、19.21%,接种AMF可明显提升镍胁迫下桂花叶片的抗氧化酶活性,活性氧清除能力有明显增加,细胞受伤害程度有明显降低,这与尹大川等的研究结果[14,19]较为一致。
另外,本研究结果表明,随Ni胁迫质量浓度的增大,桂花幼苗植株株高、茎粗、总干物质质量呈逐渐下降趋势,接种AMF的植株株高、茎粗、总干物质质量均明显高于未接种处理;Ni胁迫质量浓度为400 mg/L时,接种AMF的桂花株高、总干物质质量分别较未接种的显著增加22.12%、15.76%,这可能是由于接种AMF可明显提升镍胁迫下桂花的光合速率,缓解镍胁迫对桂花生长发育造成的伤害,进而促进桂花生长,这与尹大川等的研究结果[14,19]较为一致。
参考文献:
[1]黄益宗,郝晓伟,雷鸣,等. 重金属污染土壤修复技术及其修复实践[J]. 农业环境科学学报,2013,32(3):409-417.
[2]Mcbride M B. Environmental chemistry of soils[M]. New York:Oxford Univemity Press,1994.
[3]刘仕翔,黄益宗,罗泽娇,等. 褪黑素对水稻幼芽镍胁迫的缓解作用[J]. 环境科学,2017,38(4):1675-1682.
[4]黄锦孙,韦东普,郭雪雁,等. 田间土壤外源铜镍在小麦中的累积及其毒害研究[J]. 环境科学,2012,33(4):1369-1375.
[5]姚俊朝,殷永泉,崔兆杰,等. Ni和V胁迫对翅碱蓬和棉花生长生理的影响[J]. 环境科学与技术,2013,36(12):33-37.
[6]贾振宇,于洁,德英,等. 丛枝菌根真菌接种对羊草抗旱性的影响[J]. 干旱区资源与环境,2017,31(1):132-136.
[7]Matsubara Y,Tamura H,Harada T. Growth enhancement and verticillium wilt control by vesicular-arbuscular mycorrhizal fungus inoculation in eggplant Solanum melongena[J]. Journal of the Japanese Society for Horticultural Science,1995,64(3):555-561.
[8]Gaur A,Adholeya A. Prospects of arbuscular mycorrhizal fungi in phytoremediation of heavy metal contaminated soils[J]. Current Science,2004,86(4):528-534.
[9]Gohre V,Paszkowski U. Contribution of the arbuscular mycorrhizal symbiosis to heavy metal phytoremediation[J]. Planta,2006,223(6):1115-1122.
[10]Li X L,Christie P. Changes in soil solution Zn and pH and uptake of Zn by arbuscular mycorrhizal red clover in Zn-contaminated soil[J]. Chemosphere,2001,42(2):201-207.
[11]Hildebrandt U,Regvar M,Bothe H. Arbuscular mycorrhiza and heavy metal tolerance[J]. Phytochemistry,2007,68(1):139-146.
[12]黃晶,凌婉婷,孙艳娣,等. 丛枝菌根真菌对紫花苜蓿吸收土壤中镉和锌的影响[J]. 农业环境科学学报,2012,31(1):99-105.
[13]王凯渊,蒋园园,宋文俊,等. AM真菌与镉互作影响桑生长和无机元素吸收转运[J]. 菌物学报,2017,36(7):996-1009.
[14]尹大川,邓勋,宋小双,等. Cd胁迫下外生菌根菌对樟子松生理指标和根际土壤酶的影响[J]. 生态学杂志,2017,36(11):3072-3078.
[15]Zhang X H,Wang L M,Zhang X,et al. Effects of arbuscular mycorrhizal fungi on upland rice oxidative stress induced by Cu and Pb contamination in soil[J]. Agricultural Science & Technology,2014,15(1):123-126.
[16]巴青松,张根生,凌玉,等. 根施甜菜碱对镍胁迫下小麦幼苗生长生理的影响[J]. 西北植物学报,2017,37(2):315-320.
[17]孙天国,沙伟,张建. 水杨酸对镍胁迫下甜瓜幼苗生理活性的影响[J]. 北方园艺,2010(11):39-41.
[18]Phillips J M,Hayman D S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection[J]. Transactions of the British Mycological Society,1970,55(1):158-161.
[19]杨国,卢可,朱高樑,等. 丛枝菌根真菌摩西球囊霉对铜胁迫下白术幼苗光合特性及抗氧化酶活性的影响[J]. 植物生理学报,2018,54(4):618-626.
[20]樊瑞苹,周琴,周波,等. 盐胁迫对高羊茅生长及抗氧化系统的影响[J]. 草业学报,2012,21(1):112-117.
