低温胁迫下橄榄耐寒性研究
2019-01-03彭远琴赵金星邱志浩佘文琴
彭远琴 赵金星 邱志浩 佘文琴


摘要:以橄榄5个品种(三棱榄、梅埔甜榄、长营、惠圆、北溪檀香)叶片为试材,采用相对电渗率拟合Logistic方程测定低温半致死温度;对橄榄5个品种束缚水/自由水、丙二醛含量以及过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、过氧化物酶(POD)活性进行测定,探讨橄榄不同品种间耐寒性差异。结果表明,随着处理温度降低和处理时间延长,相对电渗率增加,且处理温度降低的叶片相对电渗率增幅比处理时间延长的大;拟合Logistic方程得到橄榄5个品种半致死温度为-2.79~-3.76 ℃;不同耐寒性的橄榄丙二醛(MDA)含量、束缚水/自由水和保护酶活性有差异。
关键词:橄榄;低温胁迫;半致死温度;Logistic方程;生理指标
中图分类号: S667.501文献标志码: A
文章编号:1002-1302(2019)21-0207-03
收稿日期:2018-08-27
基金项目:福建省科技重大专项(编号:2013NZ0002-1C)。
作者简介:彭远琴(1993—),女,重庆人,硕士研究生,主要从事园艺植物生理研究。E-mail:Pengyuanqin0@163.com。
通信作者:佘文琴,博士,教授,主要从事果树生理生化研究。E-mail:wenqinshe@163.com。
橄榄[Canarium album (Lour.) Raeusch]又名青果,为橄榄科橄榄属热带亚热带果树,果实营养十分丰富,属福建省药食两用的特色经济水果[1]。橄榄喜温暖,不耐寒冷,福建橄榄以闽侯、闽清种植最多,据闽清县气象局有关资料显示,闽清县自1991—2016年每隔8~10年就有1次较为严重的极端低温周期性冻害,导致橄榄枝叶枯干、幼树冻死、产量下降[2]。低温成为了影响橄榄正常生长和产量提高的限制性因素,因此鉴定橄榄耐寒性程度对橄榄产业发展至关重要。
目前关于橄榄耐寒性的研究多集中在耐寒相关生理指标的分析[3-6],对橄榄低温半致死温度的研究鲜有报道。本试验选用三棱榄、梅埔甜榄、长营、惠圆、北溪檀香5个橄榄品种叶片,通过采用相对电渗率配合Logistic方程研究橄榄耐受低温程度和相关生理指标来探讨5个品种间耐寒性关系,旨在为橄榄引种栽培、扩大栽培区域提供理论和实践基础。
1材料与方法
1.1试验材料
供试材料采自福建省福州市果树良种场(福州市晋安区新店镇凤池村,119°27′E、26°14′N)。以三棱榄、梅埔甜榄、长营、惠圆、北溪檀香10年实生树为试材,于2018年1月11日,每个品种选取位于同一等高线,长势、水培管理较一致的3株生物重复,每株取树冠外围东、南、西、北4个方位无病虫害春梢枝条放入保鲜盒带回。
1.2试验方法
1.2.1低温处理
带回的枝条,摘取从顶端算起第2张至第3张复叶,用自来水冲洗干净,用双蒸水洗3次后用纱布擦干水分,将同一品种的叶片随机混匀,分别置于15(CK)、1、-1、-3、-5、-7 ℃冰箱分别处理1、2、3、4、5、6、12 h,处理后的样品于室内静置30 min解冻,用于相对电渗率测定。
1.2.2电渗率测定
按照设置的时间梯度处理橄榄叶片。用圆形打孔器(直径0.5 cm)将解冻后的叶片避开主叶脉位置打孔,将小圆片快速称取0.1 g放入装有10 mL双蒸水的试管内,室内浸提12 h,重复3次。用DDS-307A电导率仪测量浸提液电渗率S1,沸水浴30 min后冷却至室温摇匀,再次测量电渗率S2,相对电渗率=S1/S2×100%。
1.2.3生理指标测定
过氧化氢酶(CAT)活性测定采用比色法;过氧化物酶(POD)活性测定采用愈创木酚法;超氧化物歧化酶(SOD)活性测定采用氮蓝四唑法;丙二醛(MDA)含量、束缚水/自由水测定参考王学奎的方法[7]。
1.3数据处理
使用Excel 2016进行数据整理和图表制作,SPSS 19.0软件进行显著性分析。对橄榄叶片相对电渗率进行非线性回归分析,用Logistic方程y=k/(1+ea-bx)拟合,式中:y为相对电渗率,%;x为处理温度,℃;k为y的最大极限值,y=k时,dy/dx=0,表示膜透性已经完全破坏;a、b为方程参数。通过线性方程y′=ln[(k-y)/y]在Excel 2016作得散点趋势图,得到a、b值和确定系数R2。x=lna/b即为半致死温度。
2结果与分析
2.1不同处理温度对橄榄叶片相对电渗率的影响
如图1所示,叶片相对电渗率随着处理温度的下降,品种间表现出不同的增幅。处理12 h,1、-1 ℃与15 ℃相比,相对电渗率上升平缓,增幅较小;-3 ℃处理后,相对电渗率迅速上升,与1 ℃相比,惠圆、北溪檀香增幅较大,分别为177.18%、151.04%,长营、三棱榄、梅埔甜榄增幅分别为42.38%、48.30%、51.38%;-3~-5 ℃处理后,橄榄叶片相对电渗率继续上升,梅埔甜榄相对电渗率增幅最大,长营次之,而北溪檀香、惠圆增幅较小;-5~-7 ℃处理后,惠圆、北溪檀香、梅埔甜榄相对电渗率上升平缓,增幅小,长营、三棱榄相对电渗率继续上升至90%左右,有较大增幅。
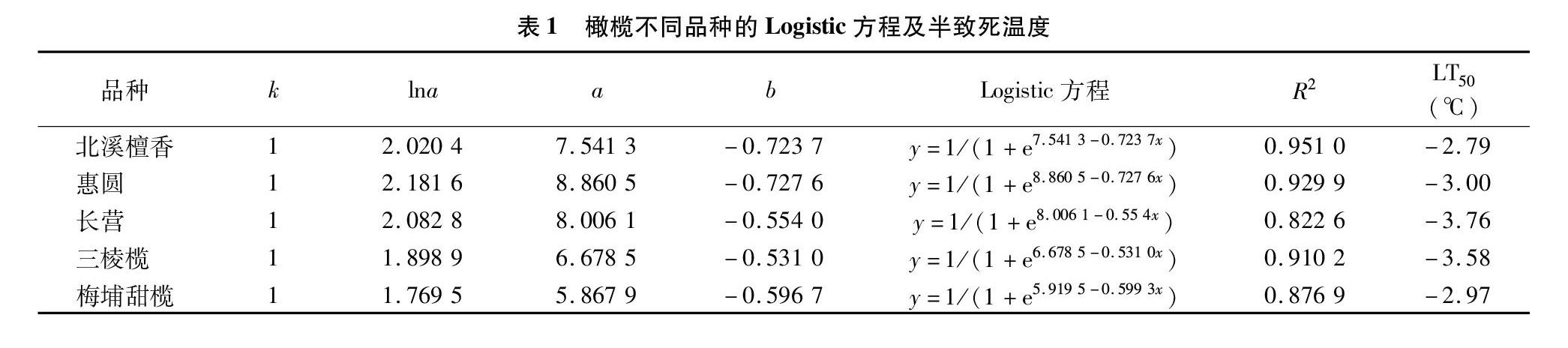
选取处理12 h的橄榄叶片相对电渗率,用Logistic方程拟合得到橄榄的半致死温度,结果见表1。由表1可以看出,橄榄5个品种的曲线方程R2在0.822 6~0.951 0之间,表明各拟合结果较精确可靠。其中,长营和三棱榄耐寒性较强,北溪檀香最弱。橄榄5个品种半致死溫度(LT50)在-2.79~-3.76 ℃ 之间。
2.2持续低温处理时间对橄榄叶片相对电渗率的影响
从表2可以看出,橄榄叶片相对电渗率随着温度降低和处理时间延长呈现不同程度的上升趋势。1、-1 ℃处理1、2、3、4、5、6 h,长营、三棱榄和梅埔甜榄与对照相比,相对电渗率上升较缓,北溪檀香、 惠圆-1 ℃处理1 h相对电渗率分别为25.73%、30.99%,处理6 h后迅速上升到41.68%、37.27%,增幅较大;-3 ℃时,随着处理时间延长,惠圆、长营、北溪檀香、三棱榄、梅埔甜榄相对电渗率上升较快,处理6 h的相对电渗率较处理1 h时分别提高20.49、17.08、34.19、9.84、1200百分点;-5 ℃处理1 h,橄榄叶片相对电渗率与对照相比上升显著,处理6 h,惠圆和北溪檀香相对电渗率超过85%;-7 ℃处理1 h,长营、三棱榄和梅埔甜榄的相对电渗率接近90%。
2.3冬季橄榄叶片保护酶活性、束缚水/自由水、丙二醛含量情况
由表3可知,橄榄各品种间保护酶活性及束缚水/自由水、MDA含量有明显差异。长营保护酶活性显著高于北溪檀香,其中长营POD、SOD活性是北溪檀香的1.64倍、1.44倍,且POD、SOD活性各个品种间总体差异显著;长营的CAT活性分别是北溪檀香、三棱榄、梅埔甜榄、惠圆的1.35倍、1.04倍、131倍、1.06 倍;北溪檀香MDA含量显著高于长营、三棱榄、梅埔甜榄和惠圆,分别是它们的1.44倍、1.27倍、1.18倍、1.23倍;长营束缚水/自由水最高,三棱榄次之,北溪檀香最低。
3讨论与结论
低温胁迫下,胞内电解质发生渗透,因此相对电渗率可以用来表征质膜受损情况,并认为相对电渗率与Logistic方程拟合得到半致死温度更能准确反映植物对低温的耐受程度[8-10]。本试验运用此法,测得15~-7 ℃处理12 h的橄榄叶片相对电渗率,与Logistic方程拟合后得到北溪檀香、惠圆、长营、三棱榄和梅埔甜榄的半致死低温分别为-2.79、-3.00、-3.76、-3.58、-2.97 ℃。其中,长营对低温耐受性更强,三棱榄次之,北溪檀香耐受性最弱,惠圆和梅埔甜榄的半致死低温差异不大;橄榄叶片相对电渗率随处理时间不同而变化,-3 ℃处理6 h与处理1 h相比,各橄榄品种叶片相对电渗率出现不同程度的增幅,说明延长低温处理时间加大了对橄榄叶片质膜伤害程度;北溪檀香、三棱榄等在-3 ℃下处理6 h的相对电渗率比在-5 ℃下处理1 h时的相对电渗率低,说明降低处理温度对质膜的伤害要比延长处理时间对质膜的伤害更大,这在低温持续处理时间方面与前人研究结果[11-13]一致。
活性氧和自由基有很强的氧化能力,对植物细胞有伤害作用。而丙二醛正是质膜受到自由基的过氧化反应产生的物质,其含量与细胞受伤害程度呈正相关,与耐寒性呈负相关[14]。CAT、POD和SOD是植物体内的保护酶,能清除活性氧和自由基,对植物细胞起保护作用,低温胁迫下,耐寒性强的品种有更高的保护酶活性[15-17]。本试验中,三棱榄、长营保护酶活性较高,丙二醛含量较低,说明耐寒性强的品种可以通过提高保护酶活性清除自由基,减少对质膜的损伤,同时减少丙二醛对质膜的2次伤害,达到保护细胞的作用,这与王旺田等的研究结果[18-19]一致;细胞内束缚水能增大原生质黏稠性,减缓代谢程度,从而增强植物的耐寒性[20],本试验中,长营、三棱榄束缚水/自由水较高,北溪檀香最低,说明植物细胞在温度变化时,耐寒性强的品种能通过增加束缚水的含量来抵御环境变化,保护植物,这在杧果[21]、梨[22]的研究上有相似结果。
综上,本试验测定半致死温度和相关生理指标得到橄榄5个品种耐寒性强弱依次为长营>三棱榄>惠圆>梅埔甜榄> 北溪檀香,在生产实践中可挑选耐寒性相对较强的品种进行种植,从而减少冻害带来的损失。
参考文献:
[1]林玉芳,陈清西. 橄榄功能成分及其抗氧化作用研究进展[J]. 热带作物学报,2010,31(1):158-163.
[2]刘义旺,黄珠英. 橄榄强霜冻害的发生类型与应对措施[J]. 福建果树,2003(3):13-14.
[3]郑家基,卢炜,陈利恒,等. 龙眼、橄榄叶片空隙率与耐寒性的关系[J]. 福建农业大学学报,1996,25(2):161-164.
[4]曾明辉,郑家基,陈利恒. 福建橄榄耐寒性快速测定法[J]. 福建果树,1999(1):13-14.
[5]韦晓霞,吴如健,胡菡青,等. 橄榄不同品种耐寒性的研究[J]. 福建农业学报,2005,20(增刊1):90-92.
[6]张小红,赵依杰,陈贻钊. 2个橄榄品种的耐寒性研究[J]. 福建林业科技,2017,44(3):90-93,98.
[7]王学奎. 植物生理生化实验原理与技术[M]. 2版. 北京:高等教育出版社,2006.
[8]郭卫东,张真真,蒋小韦,等. 低温胁迫下佛手半致死温度测定和抗寒性分析[J]. 园艺学报,2009,36(1):81-86.
[9]娄晓鸣,王化坤,陈勇明,等. 以电导法配合Logistic方程确定25个白肉枇杷的抗寒性[J]. 热带作物学报,2016,37(10):1920-1923.
[10]王玮,李红旭,赵明新,等. 7个梨品种的低温半致死温度及耐寒性评价[J]. 果树学报,2015,32(5):860-865.
[11]邢海盈. 低温胁迫下三角梅的生理变化及其调控机理的研究[D]. 福州:福建农林大学,2013:14-15.
[12]佘文琴,刘大林,齐清琳. 杨桃耐寒性初步研究[J]. 福建农林大学学报(自然科学版),2003,32(1):61-63.
[13]张天翔,曹明华,林艺华,等. 应用电导法配合Logistic方程确定“多福”甜椒的抗寒性[J]. 分子植物育种,2018,16(12):4068-4071.
[14]马德华,卢育华,庞金安. 低温对黄瓜幼苗膜脂过氧化的影响[J]. 园艺学报,1998,25(1):61-64.
[15]董万鹏,罗充,龙秀琴,等. 低温胁迫对西番莲抗寒生理指标的影响[J]. 植物生理学报,2015,51(5):771-777.
[16]佘文琴,刘星辉. 越冬期番石榴叶片脂质的过氧化状况[J]. 福建农业大学学报(自然科学版),2001,30(3):357-361.
[17]柴文臣,马蓉丽,焦彦生,等. 低温胁迫对不同辣椒品种生长及生理指标的影响[J]. 华北农学报,2010,25(2):168-171.
[18]王旺田,刘文瑜,姜寒玉,等. 低温胁迫对葡萄幼苗渗透调节物质及抗氧化酶活性的影响[J]. 中国果树,2015(1):14-17.
[19]钟鹏,刘杰,王建丽,等. 花生对低温胁迫的生理响应及抗寒性评价[J]. 核农学报,2018,32(6):1195-1202.
[20]马翠兰,刘星辉,胡又厘. 柚品种间的耐寒性差异及其机理[J]. 福建农业大学学报,1998,27(2):160-165.
[21]佘文琴,刘星辉. 低温胁迫下杧果叶片若干生理生化变化[J]. 福建农业大学学报,2001,30(2):180-184.
[22]李玉梅,陳艳秋,李莉. 梨品种枝条膜透性和水分状态与抗寒性的关系[J]. 北方果树,2005(1):3-5.
