库尔勒香梨自交不亲和SFBB-γ基因F-box区和可变区V3 RNAi表达载体的构建及遗传转化
2018-11-12钟颖冯建荣刘海楠李文慧吕文娟田雯
钟颖,冯建荣*,刘海楠,李文慧,吕文娟,田雯
(石河子大学农学院/特色果蔬栽培生理与种质资源利用兵团重点实验室,新疆 石河子 832003)
梨属(PyrusL.)属蔷薇科(Rosaceae)梨亚科(Pomoideae)[1],其品种繁多且资源非常丰富。库尔勒香梨(Pyrusbretschneideri Rehd)是新疆重点发展的特色果树品种之一[2],但库尔勒香梨自花授粉结实力低,是极具代表性的配子体型自交不亲和性(Gametophytic self-incompatibility,GSI)果树[3],生产上常采用蜜蜂授粉或人工授粉的方式来提高经济效益,这使梨园的管理变得复杂化,加大了投入成本,所以,针对其自交不亲和性机理展开深层次探究,优选培育自交亲和性香梨品种对实际生产有着重要意义。
果树自交不亲和性由S位点复等位基因控制[4],该位点的基因至少包括雌蕊决定子S-RNase基因和编码 F-box蛋白的花粉S基因[5]。现阶段以蔷薇科的果树S-RNase基因为方向的探究活动获取了诸多的成果,在李属果树中也已经确定花粉S决定子的最佳候选基因为 SLF(S locus F-box gene)/SFB(S haplotypespecific F-box gene)基因[6],梨亚科花粉 S单元型中具有诸多的同源拷贝,F-box基因家族十分庞大,目前从梨属和苹果属果树中分离鉴定出的多数为 SFBB(S locus F-box brothers)基因[7],该类基因N端有一个F-box结构域,由40-50氨基酸所聚合而成,和S-RNase基因连锁同时在花粉里呈现出特异性表达,具有S等位基因的多态性,因而在诸多的梨亚科花粉S决定子的候选基因当中被视作首选[8]。Cheng等[9]最先在苹果(Malus × domestica)花粉中克隆到了S单元型多态性基因MdSLFB1(SLFB :S-RNase linked F-box) 和 MdSLFB2;Sassa等[7]通过筛选BAC文库的方法克隆到了4个苹果SFBB 基 因(MdSFBB3-α、MdSFBB3-β、MdSFBB9-α和MdSFBB9-β)和6个梨属SFBB基因。在S位点内,S-RNase基因的两端均有梨属SFBB基因存在,一个上述位点单元里SFBB基因涵盖了诸多的序列相似性很高的基因(α、β、γ)[7]。这些基因均有S位点特性同时具有遗传多态性,然而目前尚不清楚与S-RNase基因相互作用产生自交不亲和性的到底是哪一类或哪几类SFBB基因,还需要对这些基因的结构、功能以及基因间的联系等进行更深入的研究。SFBB-γ 基因是梨属众多SFBB基因中的一种,它由一个 F-box区域和 4 个可变区(V1、V2、V3、V4)组成,目前已有许多SFBB-γ基因被分离鉴定出来[10]。
RNAi技术具有特异性、高效性、可传播性、遗传性、高稳定性和ATP依赖性[11]。这些特性保证了对目的基因的精确沉默,可用于特定基因的功能研究[12]。Jung等[13]借助RNAi技术,使油菜(B.rapa cv)的 S位点花粉识别控制基因 S60基因沉默,所得到的RNAi转基因植株自交亲和并且能够使后代具有同等特性。与 S-RNase基因有关的其余基因会对自交不亲和性造成干扰[14],RNAi降低植物自交不亲和相关因子如苹果的 Md ABCF[15]和茄属植物的 Sa CUL1(Cullin1蛋白)[16]表达,使其自交不亲和反应失效。这些研究为亲和育种提供新的方向。所以,将此技术运用于探究SI的活动当中,在果树亲和突变育种方面能够收获可喜的成效。
本研究以自交不亲和库尔勒香梨的 SFBB-γ基因为研究对象,以 SFBB-γ 基因 F-box区和可变区 V3区为干扰利用目标,利用融合 PCR构建ihpRNA(intron-containing hairpin RNA)结构,再结合酶切连接的方式构建 SFBB-γ 基因的 RNAi植物表达载体,运用冻融转化法在农杆菌GV3101里转进 SFBB-γ 基因的 RNAi植物表达载体。利用农杆菌介导的方法转化库尔勒香梨无菌苗,通过抗性筛选和 GUS化学染色法检测转化的情况。
SFBB(S-haplotype-specific F-box protein)基因的F-box区和可变区与亲和突变密切相关,本实验利用 RNAi技术,对新疆重点发展的特色果树品种‘库尔勒香梨’的花粉 F-box区和可变区 V3序列进行干扰,从而使梨自交不亲和性的调节控制得以达成且成功培育出特色梨自交亲和品种。本研究成果对特色梨自交亲和品种的培育及自交不亲和机制的研究具有重要的作用。
1 材料与方法
1.1 材料
于2016年3月末至4月初,采集新疆轮台国家果树资源圃自交不亲和库尔勒香梨成龄树气球期的花,剥取花药于阴凉干燥处晾干至散粉,收集花粉置于-70℃ 低温保存。并采集1年生休眠枝条,将休眠枝条置于室内进行浸泡催芽,用于初代外植体的培养,参照本实验室的消毒方法获得无菌苗[17],根据本实验室建立的库尔勒香梨再生体系获得再生不定芽。
GV3101农杆菌菌株(链霉素抗性,Sm)和植物表达载体 pCAMBIA1304(12361pb,卡那霉素抗性,Km;含 GUS基因)由石河子大学生命科学学院提供;克隆载体 PMD19-T、限制性内切酶、T4 DNA 连接酶、PCR产物纯化试剂盒购于TAKARA-宝生物工程(大连)有限公司;大肠杆菌菌株(DH5α)由本实验室保存;质粒提取试剂盒购于Omega生物技术公司(美国),DL 2000(Marker)、2×Taq PCR Master Mix购于天根生化科技有限公司(北京),电泳用琼脂糖、配置所需固体/液体培养基的相关药品及抗生素购于生工生物工程股份有限公司(上海)。引物合成和测序均由生工生物工程股份有限公司(上海)完成。
1.2 方法
1.2.1 序列分析及引物设计
经生物信息学分析,本实验室已获得的库尔勒香梨 SFBB-γ基因 cDNA全长序列含有 SFBB-γ基因的完整结构,即 F-box结构域、4个可变区(V1、V2、V3 和 V4),有研究表明,SFBB-γ 基因的F-box区域氨基酸的变异较多,氨基酸的变异存在于 V1,V3,V4中,而 V2中未发现[10],故 RNAi目标是选择本品种的特异性变异区,其中V1片段太短,V4尚未确定获得,故选择 F-box区和 可变区V3作为干扰目标,可借助RNAi技术加以影响从而达成梨的亲和突变。按照RNAi表达载体的建立基本准则和干扰对象,挑选 SFBB-γ基因将相应对象锁定为F-box区及可变区V3。用Primer5.0软件设计引物 F-box 区:FbP1、FbP2、FbP5、FbP6;可变区 V3:V3P1、V3P2、V3P5、V3P6(表 1)分别扩增到 SFBB-γ基因的反向和正向片段,作为RNAi发卡结构的两臂(反向互补)。因为内含子片段未出现在所得到的SFBB-γ片段里,为使沉默效率得到确保,插进棉花基因组DNA的序列(西南大学生物技术中心提供)设计引物 FbP3/FbP4;V3P3/V3P4扩增(表 1)作为间隔片段,用于构建 ihpRNA结构。通过2次PCR反应分别融合 SFBB-γ基因 F-box区和可变区V3的正向、间隔片段、反向片段基因构建 ihpRNA结构。

表1 RNAi载体构建的引物Tab.1 The primer of RNAi vector construction
1.2.2 SFBB-γ基因F-box区和可变区V3 ihpRNA的构建
引 物 FbP1、FbP2、FbP5、FbP6;V3P1、V3P2、V3P5、V3P6分别以 SFBB-γ基因3′端全长序列的菌液为模板对SFBB-γ的F-box区及V3区的正、反向片段分别进行扩增,引物 FbP3、FbP4;V3P3、V3P4用棉花基因组DNA当做模板对SFBB-γ的 F-box区及V3区的间隔片段进行扩增。反应体系为25μL标准。参照刘月霞等[18]的PCR反应程序,其中FbP1/FbP2;FbP5/FbP6退火温度为 56 ℃;V3P1/V3P2;V3P5/V3P6退火温度为 54 ℃;FbP3/FbP4;V3P3/V3P4退火温度为59℃。回收纯化所有的PCR 产物,取1 μL回收产物进行1%电泳检测,借助离心浓缩仪把其它旋转干燥浓缩5 min(2000 rpm ,45℃),对以上产物进行等摩尔混杂合并,以融合 PCR完成 SFBB-RNAi构件的融合,无需配以引物,以TaqDNA聚合酶完成各个片段的相互补充及延展,从而产生长的融合 PCR产物。融合PCR 25μL反应体系:2×TaqMasterMix10 μL,上述等摩尔混合物10 μL。反应程序与上述相同。1.2%电泳测试上述产物,对重组片段(最大片段)进行回收,待完成纯化之后与(10 μL体系)克隆载体 PMD19-T相连,连接产物转化大肠杆菌(DH5α)感受态细胞,在结束蓝白斑筛选及菌液PCR鉴别判定之后测序阳性克隆,同时比较剖析所获取的结果。正确的重组质粒命名PMD19-RNAi-SF BB-F-box;PMD19-RNAi-SFBB-V3。
1.2.3 SFBB-γ基因F-box区和可变区V3 RNAi表达载体的构建
以PstI及KpnI酶切pCAMBIA1304同时收回载体大片段、酶切PMD19-RNAi-SFBB重组质粒且将目的小片段收回。受到T4DNA连接酶的影响(10 μl标准连接体系,4℃过夜,约 12 h),把目的片段连接于表达载体 pCAMBIA1304上的启动子及终止子之间。用 PstI和 KpnI双酶切鉴定,将鉴定正确的重组质粒命名为 pCAMBIA1304-RNAi-SFBBF-box;pCAMBIA1304-RNAi-SFBB-V3。
1.2.4 SFBB-γ基因RNAi表达载体农杆菌转化
采用液氮冻融法将pCAMBIA1304-RNAi-SFBBF-box;pCAMBIA1304-RNAi-SFBB-V3分别转化到农杆菌 GV3101 感受态细胞中。2 μL(约 50 ng)重组质粒添进农杆菌感受态细胞(20 μL)之内且将其摇晃匀称,完成30 min冰浴之后,放进液氮中进行1min的速冻,立即进行28℃水浴待其融化,接着将液体培养基(不包括抗生素)800 μL YEB添入。将上述混合液置于28℃ 振荡(180 r/min)培养2-3 h后6000 r/min离心1 min,倒掉上清至残余液体100 μL左右,吸打混匀之后将其涂布含 Km(50 μg/mL)及 Sm(125 μg/mL)的 YEB 固体培养基上,平板倒置28℃恒温培养 36-48 h后,挑选乳白色抗性菌落取出,于含Km(50 μg/mL)和Sm(125 μg/mL)的 YEB 液体培养基(约 5 mL)中,28℃ 180r/min振荡培养过夜。用引物 FbP1和FbP6;V3P1及 V3P6分别完成菌液 PCR 检验,将扩增出目的条带的菌液送样测序。
1.2.5 库尔勒香梨的转化及转基因植株鉴定
把转化有目标载体的农杆菌,划线培养,挑单克隆于2 mLYEB液体培养基之内,于28℃环境内220 r/min过夜培养到达成饱和状态。挑选活化较佳的菌液在上述环境中220 r/min依据1∶100在YEB培养基内接种,过夜培养到 OD600nm=0.5的时候,对培养到预设状态的农杆菌菌液进行分装,置于50 mL离心管之内,每管分别为20 mL,在室温环境中3500 r/min离心20 min,把上清去掉,将MS液体培养基重悬添加进来,剂量为 20 mL,并加入 100μL AS(20 mg/mL),待转染时候用。在无菌环境中,剪裁香梨叶片,使其变成方形,尺寸为 0.5 cm×0.5 cm,且在已经制备好的菌液内将其置入,进行10 min的摇晃,90 r/min。平铺香梨叶片于共培养培养基1/2 MS+TDZ 1.0 mg/L+IBA 0.5 mg/L培养(25℃,2 d)。随后转至筛选培养基 1/2 MS+TDZ1.0mg/L+IBA 0.5mg/L+Kana50mg/L+Cef 400 mg/L培养(光周期16 h/8 h,光强 3000 lx,温度 25℃)中对愈伤组织和不定芽进行引诱。在7天时对侵染叶片进行GUS染色,把 GUS染液 37℃预热,于染色板每孔中加入 1 mL,取侵染的叶片及未转化香梨植株(CK)的叶片,浸入 GUS染液,染色 24 h。在37℃恒温箱内放入染色板,使其密封不可见光。而后以 75%乙醇溶液对样品进行漂洗,使叶绿素全部脱除并拍照记录。
2 结果与分析
2.1 SFBB-γ 基因 F-box区和可变区V3 ihpRNA的构建
PCR 扩增片段(其中141 bp和120 bp分别为F-box区和 V3区发卡结构的臂,其余的为由引物的‘重叠’部分构成的邻近片段的一些序列)和间隔片段(242 bp,发卡结构的环),如图1A 和图2A所示。
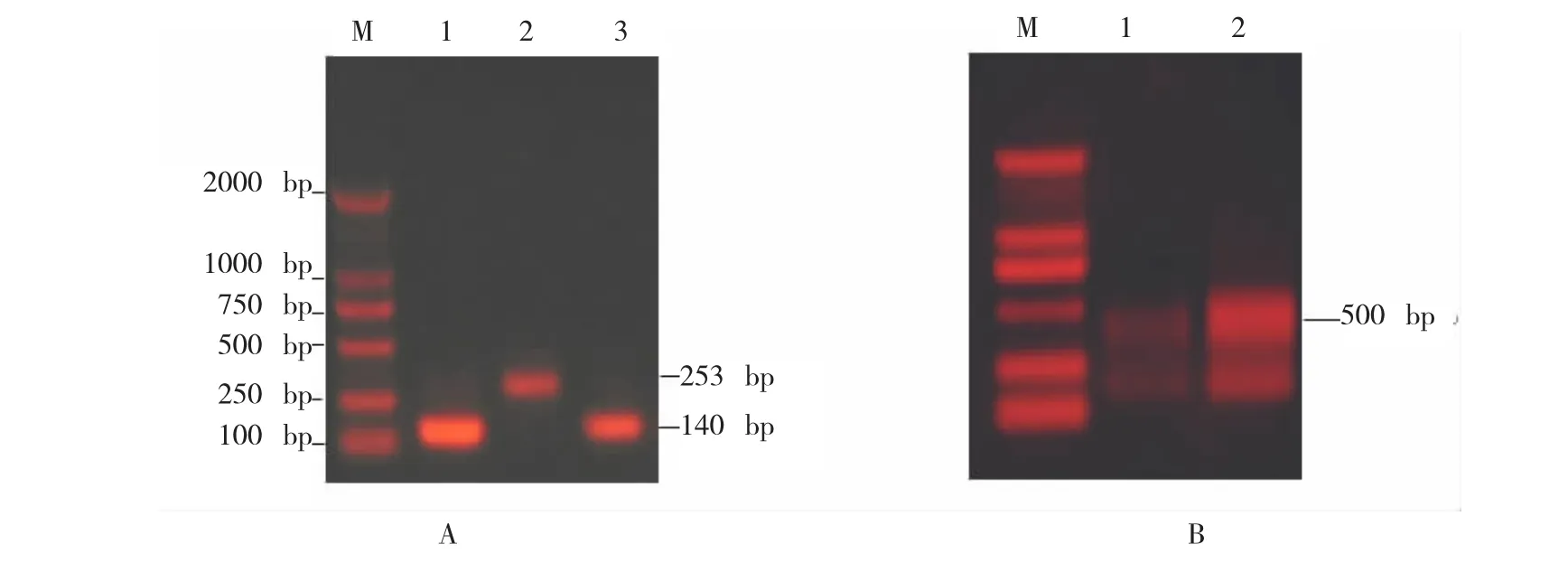

图1 Fox区正、反向和间隔片段的扩增与融合PCRFig.1 The Fox PCR amplification of Sense,antisense,interval fragment and fusion PCR

图2 可变V3区正、反向和间隔片段的扩增与融合PCRFig.2 The V3 PCR amplification of Sense,antisense,interval fragment and fusion PCR
回收浓缩后进行第二轮 PCR,得到的最大片段即为 SFBB-γ基因的 RNAi构件,其片段大小为524 bp(F-box 区)和 482 bp(V3区),如图 1B和图2B所示,PCR产物还包含有非重组结构片段(300 bp以下),只回收重组结构(最大的片段)并与PMD19-T载体连接,对阳性克隆进行测序,测序结果如图 1C和图 2C,表明片段融合成功。重组质粒为PMD19-RNAi-SFBB-F-box和PMD19-RNAi-SFBB-V3。
2.2 SFBB-γ基因F-box区和可变区V3 RNAi表达载体构建与鉴定
重组质粒 PMD19-RNAi-SFBB经 PstI和 KpnI酶切后回收小片段(含 ihpRNA结构),载体质粒PMD19-T经 PstI和 KpnI酶切后回收大片段,在T4DNA连接酶的作用下,将 RNAi构件连接到植物表达载体上,构建 SFBB-γ 基因 RNAi植物表达载体 pCAMBIA1304(图 3)。
用 PstI和 Kpn I酶切进行验证,分别得到约12700 bp和 500 bp左右(含发卡结构序列)的片段(图 4A),表明 SFBB-γ 基因的 ihpRNA 片段已经成功连接到表达载体上。

图3 RNAi表达载体示意图Fig.3 pCAMBIA 1304-RNAi-SFBB vector map
2.3 农杆菌中 SFBB-γ 基因 RNAi表达载体的菌液 PCR鉴定
液氮冻融法将 pCAMBIA1304-RNAi-SFBB转化至农杆菌 GV3101感受态细胞,在含上述 2种抗生素(Km 和 Sm)的 YEB固体培养基上筛选转化子。挑取阳性菌落培养,用 FbP1和 FbP6;V3P1和V3P6分别进行菌液 PCR扩增验证,得到与预期大小相符的片段(图4B),测序结果也与克隆载体上的 ihpRNA结构序列一致,证明表达载体已转入农杆菌 GV3101中。

图4 酶切鉴定及农杆菌转化菌液PCR验证Fig.4 Restriction identification and A.tumefaciens bacteria liquid PCR verification
2.4 叶盘转化法验证
选取未经过侵染的香梨无菌叶片(CK)和侵染的库尔勒香梨无菌苗再生苗的叶片进行GUS组织化学染色。酒精将其完全脱色后,观察染色情况,发现叶片的切伤口处和叶脉处有GUS染液的特异蓝色(图 5、图 6),说明 SFBB 基因的 F-box区和可变区V3 RNAi表达载体基因已经成功整合到了库尔勒香梨无菌苗基因组上,并且能够表达。

图5 SFBB-F-box区RNAi库尔勒香梨无菌苗转基因植株的GUS活性检测Fig.5 GUS activity detection of SFBB-F-box region RNAi transgenic tobacco

图6 SFBB-V3基因RNAi库尔勒香梨无菌苗转基因植株的GUS活性检测Fig.6 GUS activity detection of SFBB-V3 gene RNAi transgenic tobacco
3 讨论
在以融合原理为根基对ihpRNA结构进行建立的时候,基因自身的内含子序列(模板为 DNA)发挥着主要的功效[19-21]。内含子区不存在于本课题组所得到的SFBB-γ基因片段之内,所选棉花基因组DNA片段系陆地棉J14基因组A03亚基因组GhHSD1(11-β-hydroxysteroid dehydrogenase ,11-β-羟基类固醇脱氢酶)基因的内含子序列(81933396-81933638,242 bp),在剖析比较序列后发现,棉花序列和已知梨的基因序列的源头有异,因而可视作发卡结构的间隔区[22]。本次研究活动是以不存在内含子为前提而开展的,完成了3个片段的融合,且顺利建成了发卡结构。
与其余物种相比较,梨的遗传转化研究所获取的突破性成果较少,并且遗传转化成效偏低。由于其难以再生,加之基因型的不一样也会造成比较大的区别,再开展多项操作譬如侵染、筛养等之后,再生不定芽的概率更低。在转化完成后,多数果树(尤其是木本果树)难以获得转基因植株,无法进行后续的基因功能研究,多数研究多停滞或仍在此阶段摸索。转基因的成功与否受到多个条件的影响,其中多数研究者认为最重要的因素是:菌株的侵染力和植物材料的类型、生理状态。由于外植体的生理状态会影响叶片再生率,同时对农杆菌会呈现不同的耐受力,所以研究中采用叶盘转化法时,选取的是试管苗上部完全展开的健壮叶片作受体,推测其拥有更高的再生能力。但本试验仅获得叶片 GUS染色结果并未获得转基因再生植株,推测可能是由于用 GV3101做菌株,孟更[22]提出在梨的遗传转化试验中用 GV3101做菌株转化率几乎为零,未得到想要的结果。所以获得转基因再生植株是本试验室下一步工作重点之一。
4 结论
已知库尔勒香梨自交不亲和SFBB-γ基因cDNA全长序列是本次研究活动开展的前提,利用融合PCR分别构建臂长141 bp、茎环253 bp的SFBB-γ基因的F-box区的ihpRNA结构和臂长120 bp、茎环253 bp的SFBB-γ 基因的可变区 V3的ihpRNA结构,结合酶切连接成功构建 SFBB基因的F-box区和可变区V3两个区域的RNAi表达载体。GUS组织化学染色结果表明,库尔勒香梨无菌苗叶片切伤处以及叶脉处有特异蓝色,说明SFBB基因的F-box区和可变区V3目的基因已成功转化到库尔勒香梨无菌苗的基因组中并且能够表达。为诱导香梨 SFBB-γ基因转录后基因沉默、获得自交亲和的梨提供参考。
