血 HLA-A、-B、-C、-DRB1、-DQB1 等位基因多态性与肝细胞癌发生关联研究*
2018-11-09欧阳石陈小平程涛钟宋刘崇文肖露露陈阳述
欧阳石,陈小平,程涛,钟宋,刘崇文,肖露露,陈阳述
HBV感染呈世界性流行,但不同地区HBV感染的流行强度差异很大。据世界卫生组织报道,全球约20亿人曾感染过HBV,其中2.4亿人为慢性HBV感染者,每年约有65万人死于HBV感染所致的肝功能衰竭、肝硬化和肝细胞癌(HCC)[1]。在全球肝硬化和HCC患者中,由HBV感染引起的比例分别为30%和45%[2]。在我国肝硬化和HCC患者中,由HBV感染引起的比例分别为60%和80%[3]。2006年全国乙型肝炎血清流行病学调查表明,1~59岁一般人群HBsAg携带率为7.18%[4-6]。据此推算,我国有慢性HBV 感染者约9300万人[7-9]。由于乙肝疫苗接种的普及,HBV感染明显减少。台北的一项研究对8733名在1987年7月后出生的高中生进行检测,发现HBsAg阳性率为1.9%,同时抗HBs阳性率仅为48.3%,提示有相当部分完成免疫接种者仍会丧失对HBsAg的免疫记忆[10]。遗传易感性是HCC发生的主要危险因素之一,在乙型肝炎发展为HCC的过程中起着关键性的作用[11]。人类白细胞抗原(human leukocyte antigen,HLA)最显著的特征是具有明显的多态性,其多态性的差异决定免疫反应的不同。由于不同个体HLA等位基因的差异,表达不同的HLA抗原,对同一抗原的免疫应答强度也不同,进而导致疾病转归不同。因此,携带不同HLA基因个体对HBV感染的易感性、发展为慢性乙型肝炎(chronic hepatitis B,CHB)和HCC发生均存在个体差异[11]。有关HLA基因多态性与HCC发生相关性的研究已经有一些报道,由于这些研究所选择的人种和地域不同,得出的结论也存在差异。本研究采用聚合酶链反应-直接测序分型(polymerase chain reaction sequence-based typing,PCR-SBT)法对广东地区HCC患者血HLA-A、-B、-C、-DRB1、-DQB1基因多态性进行了分析,为从基因水平研究免疫遗传因素在汉族人群HCC发病机制中的作用提供参考。
1 资料与方法
1.1 研究对象 2012年6月~2015年12月 就诊于解放军第458医院的广州地区HBV感染后罹患HCC患者56例,男性53例,女性3例;年龄(51.38±10.21)岁。符合中华医学会制定的原发性肝癌诊疗规范标准[12],排除合并HCV或/和HIV感染者。另选在深圳血库义务献血的健康志愿捐献者97例,男性86例,女性11例;年龄(49.71±12.34)岁,两组间年龄和性别均无统计学差异(P>0.05)。
1.2 血清学检测 采用ELISA法检测HBsAg(北京万泰生物药业有限公司);采用PCR法检测HBV DNA(Light Cycler@480荧光定量 PCR仪和LightCycl er@480 SYBRGreen I Master试剂盒)。
1.3 血液HLA基因分型 抽取静脉血,采用EDTA抗凝,抽取0.3 mL,使用Gentra DNA提取试剂(美国,Gentra)抽提DNA。采用 SBT法(美国Ariza公司试剂盒,即Atra Genetics AlleleSEQR HLA-A,-B、-C、-DQB1、-DRB1 SBT)分别扩增 HLA-A、B 等位基因第2、3和4外显子、HLA-C和-DQB1基因第2和第3外显子、HLA-DRB1基因第2外显子,纯化产物后,进行测序反应,使用ABI3730型序列分析仪进行毛细管电泳。应用Assign version 3.5版(Conexio Genomics,AppIecross,澳大利亚)软件分析测序图谱。
1.4 统计学方法 应用Arlequin统计软件分析基因频率,计算研究对象各检测位点等位基因和基因型频率,并验证是否符合Hardy-Weinberg平衡,此处以P<0.05作为Hardy-Weinberg不平衡的标准。应用Pearson卡方检验或连续校正卡方检验或Fisher精确概率法计算两组人群基因频率差异,以P<0.05为差异具有统计学意义。
2 结果
2.1 HCC患者与健康人血HLA-A等位基因的情况 HCC患者血HLA-A*02:03和A*02:07等位基因频率明显高于健康人(x2=5.167、x2=10.33,P=0.023、P=0.001);HCC 患者血 HLA-A*30:01等 位基因频率0.017857明显低于健康人(x2=5.355,P=0.021,表 1)。
2.2 HCC患者与健康人血HLA-B等位基因频率比较 HCC患者HLA-B*46:01等位基因频率明显高于健康人(x2=5.439,P=0.02);HCC 患者 HLAB*13:02等位基因频率明显低于健康人(x2=6.702,P=0.01,表 2)。
2.3 HCC患者和健康人HLA-C等位基因频率分析 两组之间未见有明显统计学差异的等位基因频率,见表3。
2.4 HCC患者和健康人血HLA-DRB1等位基因频率分析 HCC患者血HLA-DRB1*14:54等位基因频率明显高于健康人(x2=14.502,P=0.000);HCC 患者血HLA-DRB1*12:02等位基因频率明显高于健康人(x2=4.761,P=0.029,表 4)。
2.5 HCC患者和健康人血HLA~DQB1等位基因频率分析 HCC患者血HLA-DQB1*05:03等位基因频率明显高于健康人(x2=4.951,P=0.026,表 5)。

表1 HCC患者与健康人血HLA-A等位基因频率比较

表2 HCC患者和健康人HLA-B等位基因频数比较

表2 -续表

表3 HCC患者和健康人血HLA-C等位基因频数比较
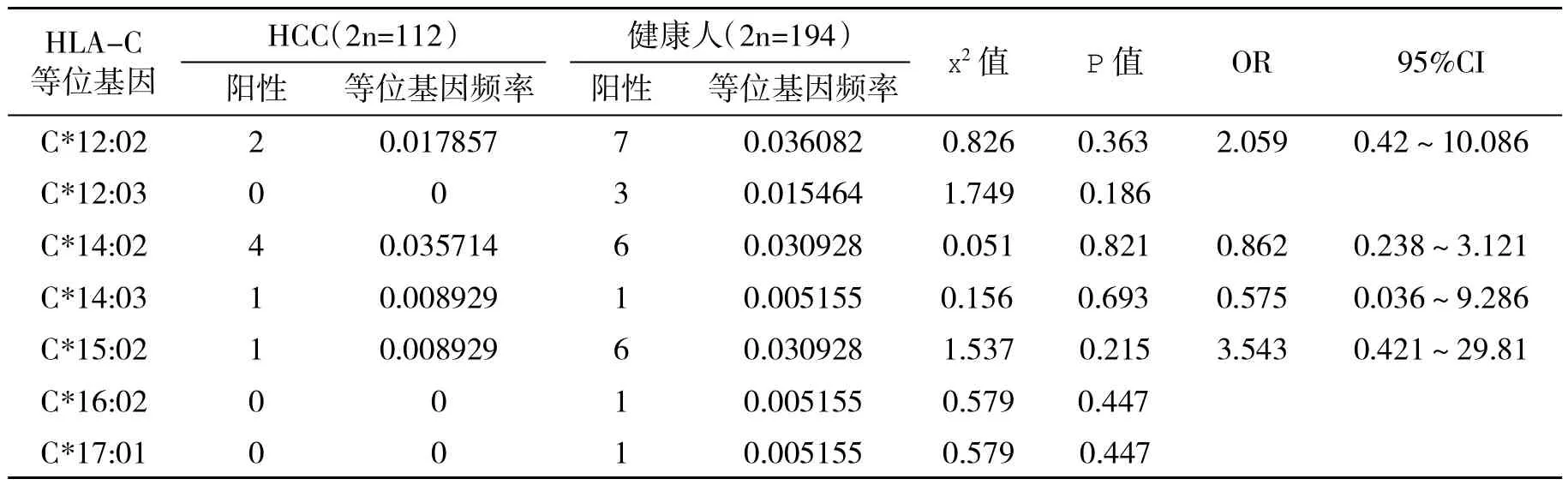
表3 -续表
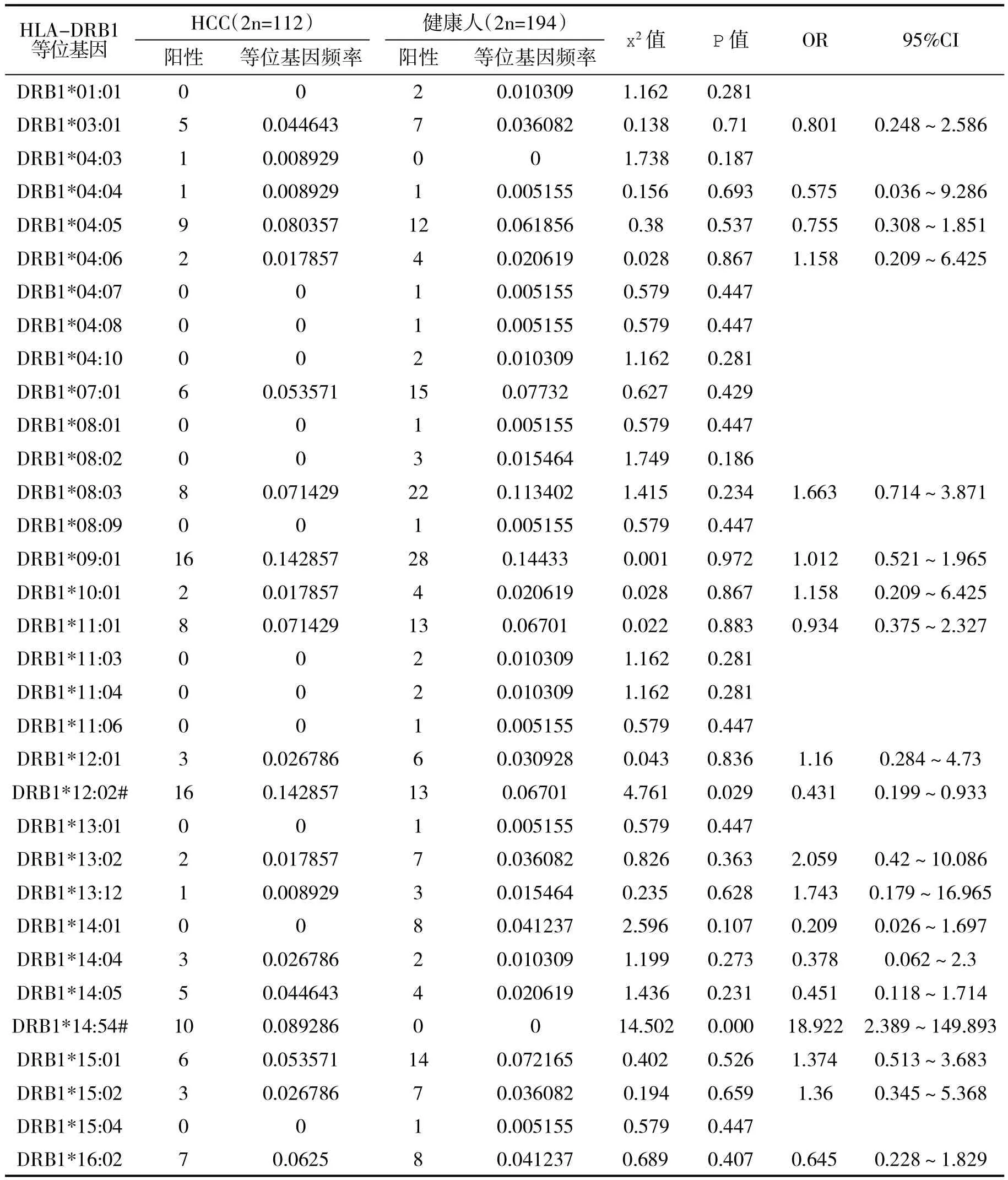
表4 HCC患者与正常人血HLA-DRB1等位基因频数比较

表5 HCC患者与正常人血HLA-DQB1等位基因频数比较
3 讨论
SBT技术无论在分型精确度、工作效率和自动化程度方面都明显优于其它分型方法。在本研究中,我们采用SBT法首次对中国广东地区HCC患者血 HLA-A、-B、-C、-DRB1、-DQB1 高分辨等位基因进行了分型。
HLA基因复合体是迄今所知的人类最具有多态性的基因系统,依据编码分子的不同特性而分成3类基因区,分别称为Ⅰ类、Ⅱ类和Ⅲ类基因。研究表明,肿瘤发生与机体免疫监控失调有关,HLA I类抗原与肿瘤的发生发展关系密切。经典的HLA I类分子包括HLA-A、B、C,几乎分布于所有的有核细胞表面,是CD8+T淋巴细胞识别标志之一。HLA I类分子具有重要的免疫生物学功能:一方面HLA I类分子直接参与抗原递呈细胞(antigen presenting cell,APC)对内源性抗原的加工和处理;另一方面,在T细胞抗原识别受体(T cell receptor,TCR)特异性识别APC递呈的抗原肽过程中,必须同时识别抗原肽以及与抗原肽结合成复合物的HLA分子,才能产生激活T细胞的信号。HLA复合体通过参与抗原递呈、制约免疫细胞间的相互作用、控制机体免疫应答的发生及强度等多个方面,参与免疫调节。尽管有在HCC组织和体外培养细胞中HLAⅠ类抗原表达增多的报道[13],有关HLAⅠ等位基因与HCC发生的关系的研究报道非常少。Pan N et al在南京对中国东南地区人群的研究显示[14]:A/11:01:01G、B/35:01:01G、B/58:01:01G和 C/03:02:01G是 HCC发生的易感基因,且B/35:01:01G显示出12.04倍的HCC发生风险,与本研究发现并不一致。我们发现HLA-A*02:03、A*02:07、HLA-B*46:01 三个HLA-Ⅰ类等位基因在HCC人群表达远高于正常健康人,提示 HLA-A*02:03、A*02:07、HLA-B*46:01 可能为HBV感染后罹患HCC的易感基因,这种差异可能与地区间的人群差异有关。
HLA II类基因的等位基因多态性导致了抗原结合槽及提呈抗原肽给T细胞的效率不同而决定着不同个体对免疫应答的差异。在II类区域中又以HLA-DRB1等位基因多态性最复杂,且是机体免疫基因(Ir)所在区域,故与免疫应答关系最为密切,并与许多疾病的遗传易感性[15-19]相关。迄今,HLA II类等位基因与HCC病理以及遗传易感性的关系已有较多阐明,本研究显示,DRB1*14:54在HCC人群表达远高于正常健康人,与之前的研究结果相符[20-22],提示其为HBV感染后罹患HCC的易感基因,是乙型肝炎患者发展为HCC的危险因素,而DRB1*12:02在HCC人群的表达显著高于正常对照,与Lin的研究也是符合的,可能是HCC的保护性因素。
