湖北不同种源红椿天然居群的种实表型变异
2018-11-08蔡京勇陈文学付翠林
蔡京勇, 陈文学, 汪 洋, 付翠林, 钟 雄
(1.湖北生态工程职业技术学院,湖北武汉 430200; 2.竹山县林业局,湖北竹山 442200; 3.巴东县野三关林业站,湖北巴东 444324)
遗传多样性是一个物种生存或者适应所生存环境和发展乃至于不断进化提高的重要前提[1],对于任何一个物种来说,面对不断变化的生长环境、扩展分布及开拓新环境,就意味着其需要强大的适应力,所以该物种的遗传多样性就需要高丰富度[2]。引起物种表型变异正是遗传多样性的表现,其根源来自于多种内在及外在环境因子的影响。表型性状即物种的数量性状,受许多基因控制[3]。因此,基于表型性状来研究物种的变异特征是一种有效的方法。从19世纪达尔文进化论诞生以来,研究遗传多样性最常见的技术方法就是通过测定植株的表型性状来实现[4]。目前,对于植物表型变异的研究主要集中在2个方面,一是对于重要的经济性状,侧重于遗传分析,寻找具有优良性状的遗传材料,为遗传改良奠定基础。二是在生态上较重要的性状,主要从地理环境上开展相关研究,注重探究物种的适应性[5-6]。
红椿(ToonaciliataRoem.)为楝科(Meliaceae)香椿属(Toona)落叶大乔木。红椿材质优良,是珍贵用材树种,具有很高的经济价值[7]。红椿已被列为国家Ⅱ级重点保护野生濒危乡土树种。该树种具有良好的开发利用前景,但自然资源稀少,遗传资源尚不明确,且人为破坏严重。因此,红椿种质资源保护迫在眉睫。保护的前提是了解其居群规模和遗传背景[8],根据其现存居群大小与遗传多样性水平进行针对性的科学保护[9]。
研究一个物种不同种源的表型变异,有助于全面准确地了解不同种源遗传变异规律和种源内个体的形态变异,为优良种源的选育和种质创新提供理论依据[10]。种实是遗传的集中点[11],种实变异是遗传变异的重要特征之一[12]。植物形态主要受遗传控制,但在不同分布区也会由于适应不同生境而产生分化,其在大尺度上的变化格局通常与气候及纬度梯度有关[13]。因此,从红椿种实形态性状来研究其表型遗传变异,能在一定程度上地更好揭示其遗传变异的大小,以及一定地理区域内,在气候、经纬度梯度上的遗传多样性变化规律,对于红椿种源选择与评价,种质资源保护及利用具有重要的理论和实践意义。
1 材料与方法
1.1 研究地概况
2013—2015年对湖北不同种源红椿天然林种质资源进行全面调查。选取其中16个尺度合适的红椿天然居群设立样方,对样方内所有乔木、灌木和草木进行每木检尺。在前期研究基础上,于2015年采集种实。由于红椿结实存在大小年现象,在16个天然居群中,仅有8个居群结实良好,且基本覆盖红椿在湖北的天然分布范围。其中利川堡上村居群、宣恩金盆村居群、宣恩大卧龙居群、咸丰村木田居群属于鄂西南地区,谷城玛瑙观居群和竹山洪坪居群属于鄂西北地区,黄石黄荆山居群和通山九宫山居群属于鄂东南地区。采集种实的居群地理位置及生态因子见表1。
1.2 种实表型性状选择与测量
红椿果实采集于果实成熟期(湖北地区因纬度差异从南到北为11月上旬至12月上旬)。不同居群的结实单株不等(利川堡上村2株、宣恩金盆村4株、宣恩大卧龙5株、咸丰村木田7株,通山九宫山5株、黄石黄荆山6株、竹山洪坪3株、谷城玛瑙观5株),在8个居群37个不同单株上采集种实,每个单株上收集60个种实。各居群内取样植株间距在35 m以上。尽量选择朝南方向处于亚优势地位,树冠中部生长正常的,无严重缺陷,无明显病虫害的果实,以避免不同生长环境造成的统计差异。

表1 不同种源红椿采样群体的地理位置及生态因子
注:气象数据来源于国家气象信息中心,平均气温参考采种点附近气象站的观测数据计算获得。其他数据采用ArcGIS 10.2从WorldClim全球数据库(www.worldclim.org)提取相应气候因子。
选择相对稳定、易于获得和测量的表型性状进行分析,如果实质量、果实纵径、果实横径、种子质量、种子纵径和种子横径等。采集的果实经自然风干后,根据百粒四分法标准,每单株随机抽取30个果实[6],用电子游标卡尺(精度0.01 mm)测量每个果实的纵径和横径,并用1/1 000电子天平分别称取每个果实的质量(精度为0.01 g),重复5次。果实性状测后,剥离果壳,按照同样方法分别测定种子纵径、种子横径、种子厚度、单粒种子质量,重复5次。种实测量指标包括果纵径、果横径、单果质量、单果种子数、单种质量、种纵径、种横径、种厚度等,并计算果形指数(果纵径/果横径)、果大小指数(果纵径×果横径)、种形指数(种纵径/种横径)、种大小指数(种纵径×种横径)[6,14]。
1.3 统计分析方法
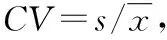
2 结果与分析
2.1 种实表型性状居群间差异
由表2可知,不同居群间红椿种实的12个指标均呈极显著差异(P<0.01),其差异显著程度由大到小依次为果纵径、果形指数、单种质量、果大小指数、单果质量、果横径、种纵径、种大小指数、种形指数、种子数、种横径、种厚度,其中果纵径、果形指数和单种质量是差异性较为突出的性状。
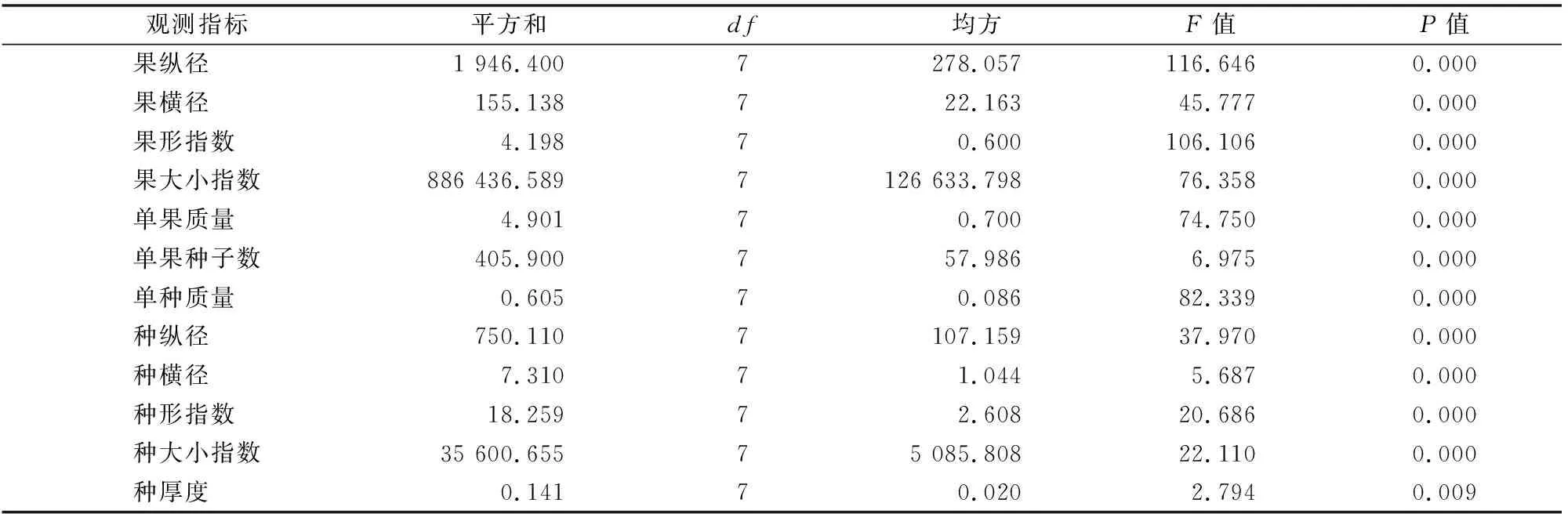
表2 湖北不同居群间红椿种实表型单因素方差分析
2.2 红椿种实表型特征分析
从表3可以看出,不同居群内不同指标值变化范围为 1.97%~18.86%。黄石居群变异系数平均值最大,竹山洪坪最小,可能与其地理区位或气候条件有关。居群内各测量或形状指标变异系数平均值均小于居群间,说明变异动力来源于不同种源的地理或环境因子。果形指数变异系数平均值最小,为3.504%,果形指数为果纵径和果横径的比值,说明其果型变异幅度最小,较为稳定。单种质量变异系数平均值最大,为13.366%,单种质量保证种子结实质量,是种源选择考虑的重要依据。
2.3 湖北不同种源红椿种实的表型多样性指数
由表4可知,8个居群种实表型Shannon-Wiener多样性指数H变化范围为1.161~1.305,Simpson多样性指数D变化范围为0.530~0.596,说明种群间存在一定的多样性,其中通山九宫山居群Shannon-Wiener多样性指数和Simpson多样性指数最高,宣恩大卧龙的Shannon-Wiener多样性指数最低,竹山洪坪的Simpson多样性指数最低。通山九宫山、黄石黄荆山和谷城玛瑙观的Shannon-Wiener多样性指数和Simpson多样性指数均呈现较高水平,变化趋势整体表现为从西部到东部逐渐升高。说明湖北不同种源红椿种实表型随地理经度的升高,多样性增大,变异相对丰富,地理变异趋势较为明显,这与居群间差异和表型特征分析结果基本一致。
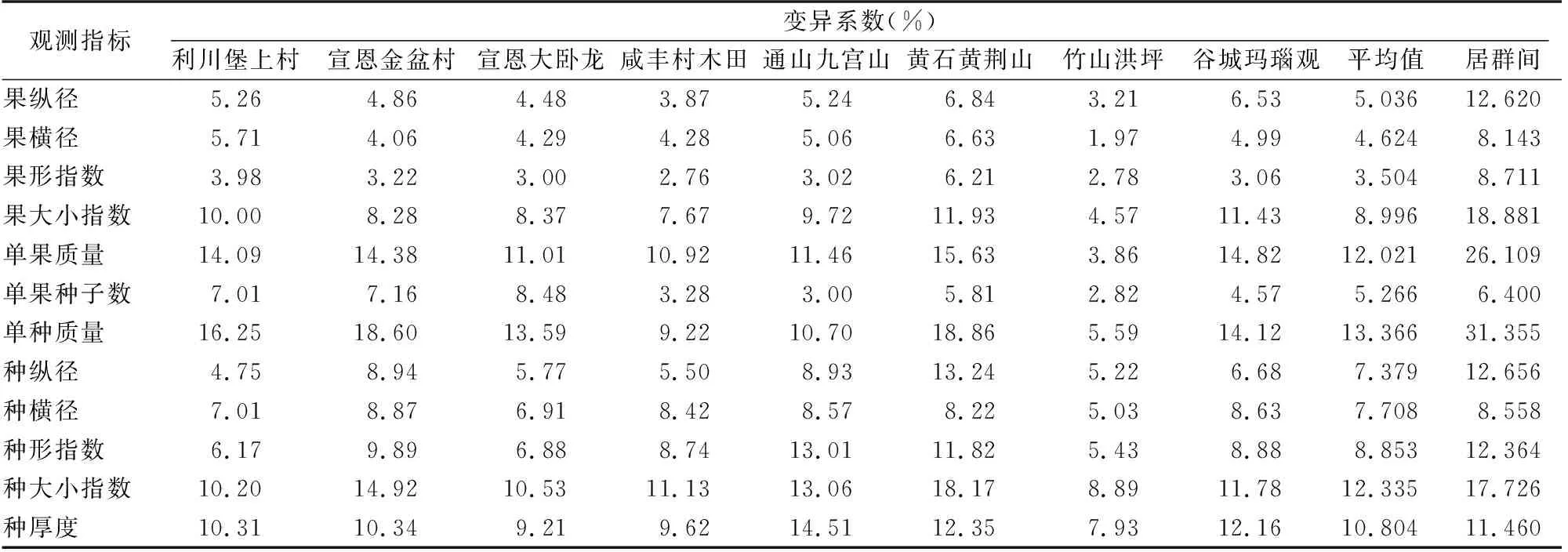
表3 湖北红椿8个居群种实表型性状变异系数
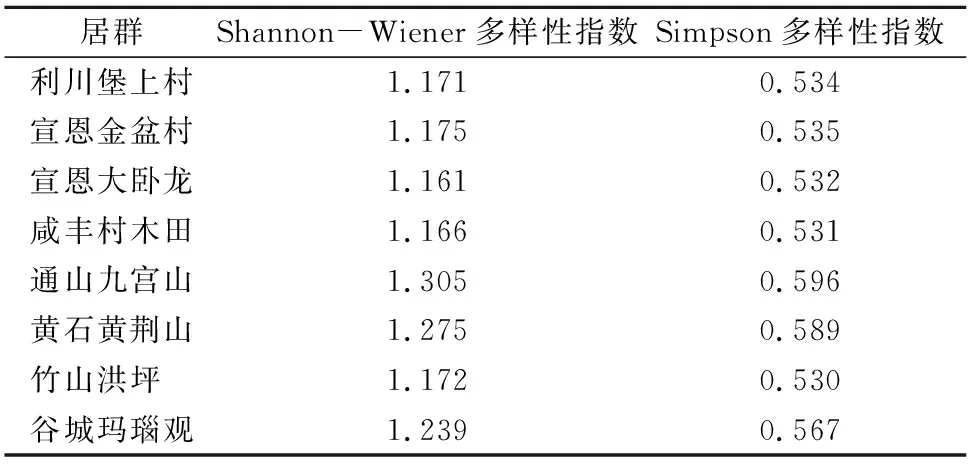
表4 湖北红椿8个居群种实表型多样性指数
2.4 种实表型性状间及其与地理环境相关性分析
表5可知,湖北红椿居群间大多数种实表型性状间显著相关。果纵径与果形指数、果大小指数、单果质量、单种质量、种纵径、种形指数、种大小指数均存在极显著正相关关系(P<0.01),与果横径呈显著相关关系(P<0.05),与单果种子数、种横径、种厚度相关性不显著。果横径与果大小指数、单果质量、单种质量呈极显著相关关系(P<0.01)。果形指数与种纵径、种形指数、种大小指数呈极显著相关关系(P<0.01)。果大小指数与单果质量、单种质量、种大小指数呈极显著相关关系(P<0.01),与种纵径呈显著相关关系(P<0.05)。单果质量与单种质量呈极显著相关关系(P<0.01),与种纵径、种形指数、种大小指数呈显著相关(P<0.05)。
由表6可知,红椿果纵径、果横径、果大小指数、单果质量、单种质量与经度呈极显著负相关关系(P<0.01)。果横径与海拔呈显著正相关关系(P<0.05)。果纵径、种纵径、种横径与无霜期呈显著正相关关系(P<0.05),种大小指数与无霜期呈极显著正相关关系(P<0.01),说明无霜期对种实形状的影响较大。年均无霜期与红椿种质资源分布有密切关系,而年积温(不是年平均温度)效应影响红椿种实的变异,同时影响红椿地理种源的分布。
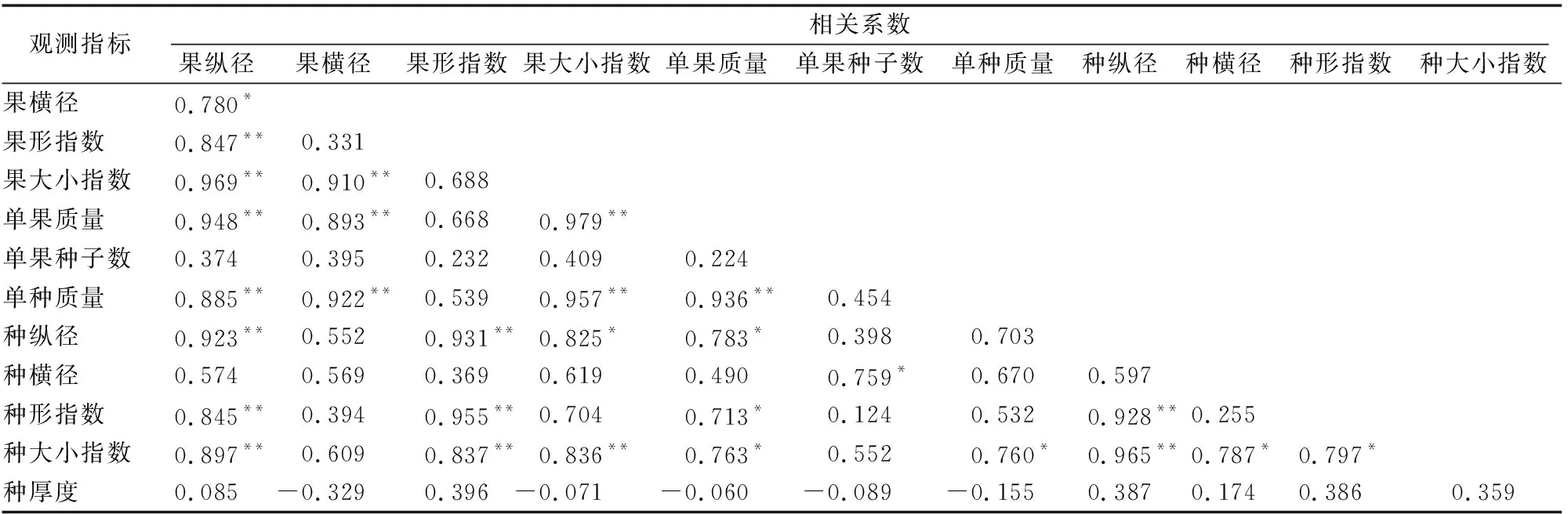
表5 湖北红椿12个种实表型性状间相关性
注:“*”表示不同表型性状间呈显著相关(P<0.05);“**”表示不同表型性状间呈极显著相关(P<0.01)。
2.5 湖北不同种源红椿居群分类
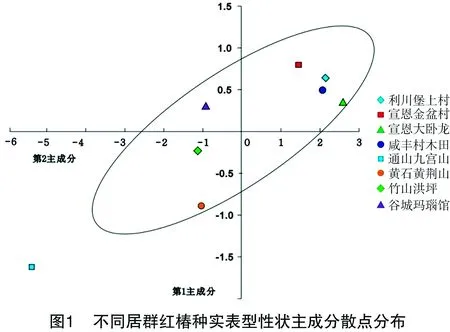
2.5.1 表型性状间的主成分分析 提取累积贡献率大于80%的前m个主成分,由表7可知,当m=2时,12个主成分累计贡献率达83.301%(>80%),说明前2个主成分可表征全部因子所代表信息,具有很高可信度。根据前2个主成分值,计算并作出主成分散点分析图,由图1可知,通山九宫山居群种实表型性状明显游离于其他居群之外。黄石黄荆山、竹山洪坪和通山九宫山居群均位于第3象限,与位于鄂西南居群和谷城玛瑙观居群离散较明显。在湖北8个天然居群中,位于鄂东南居群与位于鄂西南的居群均保持较为独立的变异连续性,呈现地理隔离模式。
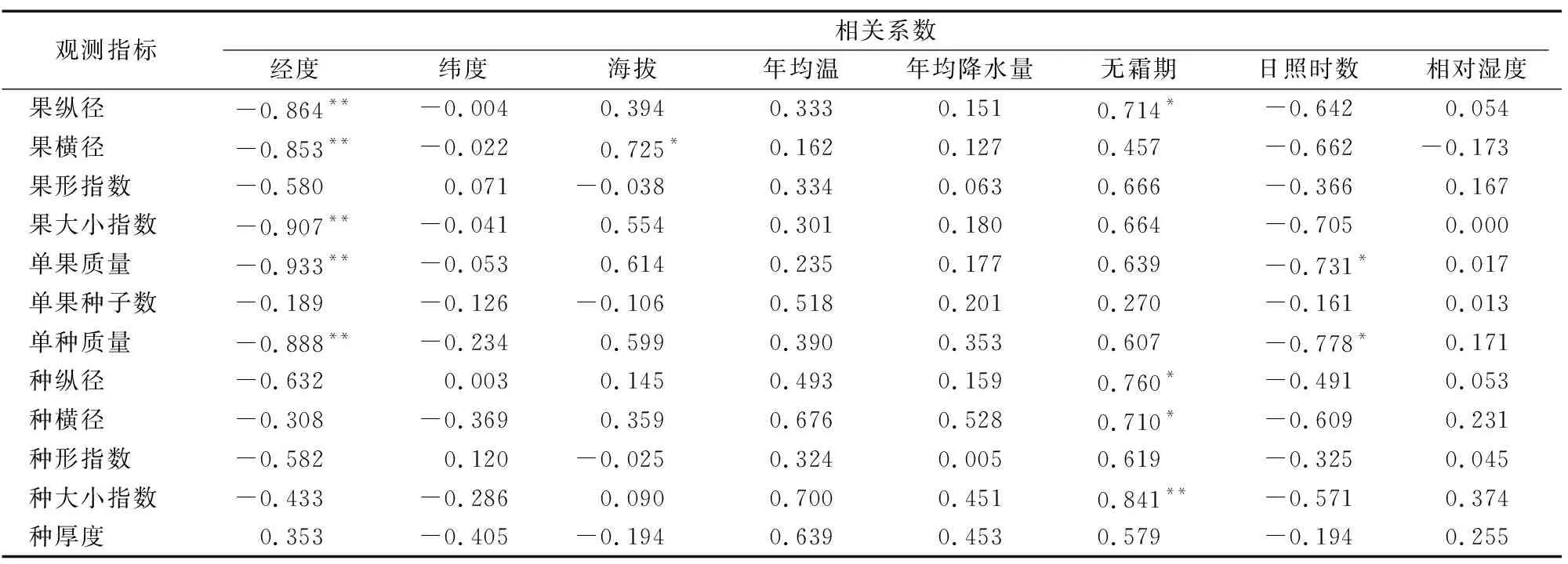
表6 湖北红椿种实表型性状与地理环境因子间相关关系
注:“*”表示果型性状与地理环境因子间呈显著相关(P<0.05),“**”表示果型性状与地理环境因子间呈极显著相关(P<0.01)。

表7 前2个主成分值、特征根、贡献率和累计贡献率
2.5.2 表型性状间的聚类分析 采用非加权平均法(UPGMA)对湖北江椿8个天然居群种实进行聚类分析。由图2可知,第1个节点为阈值δ=138.47,将谷城玛瑙观、通山九宫山、黄石黄荆山分为一个类群,利川堡上村、咸丰村木田、宣恩金盆村、宣恩大卧龙和竹山洪坪划分为一个类群。第2个节点阈值δ=100.388,将通山九宫山分为一个类群,黄石黄荆山和谷城玛瑙观聚为一个类群。依次在低节点阈值δ=58.847 时,向下划分,将咸丰村木田、宣恩金盆村、宣恩大卧龙和利川堡上村聚为一个类群,竹山洪坪居群单独为一个类群。聚类分析结果与主成分分析结果基本一致。

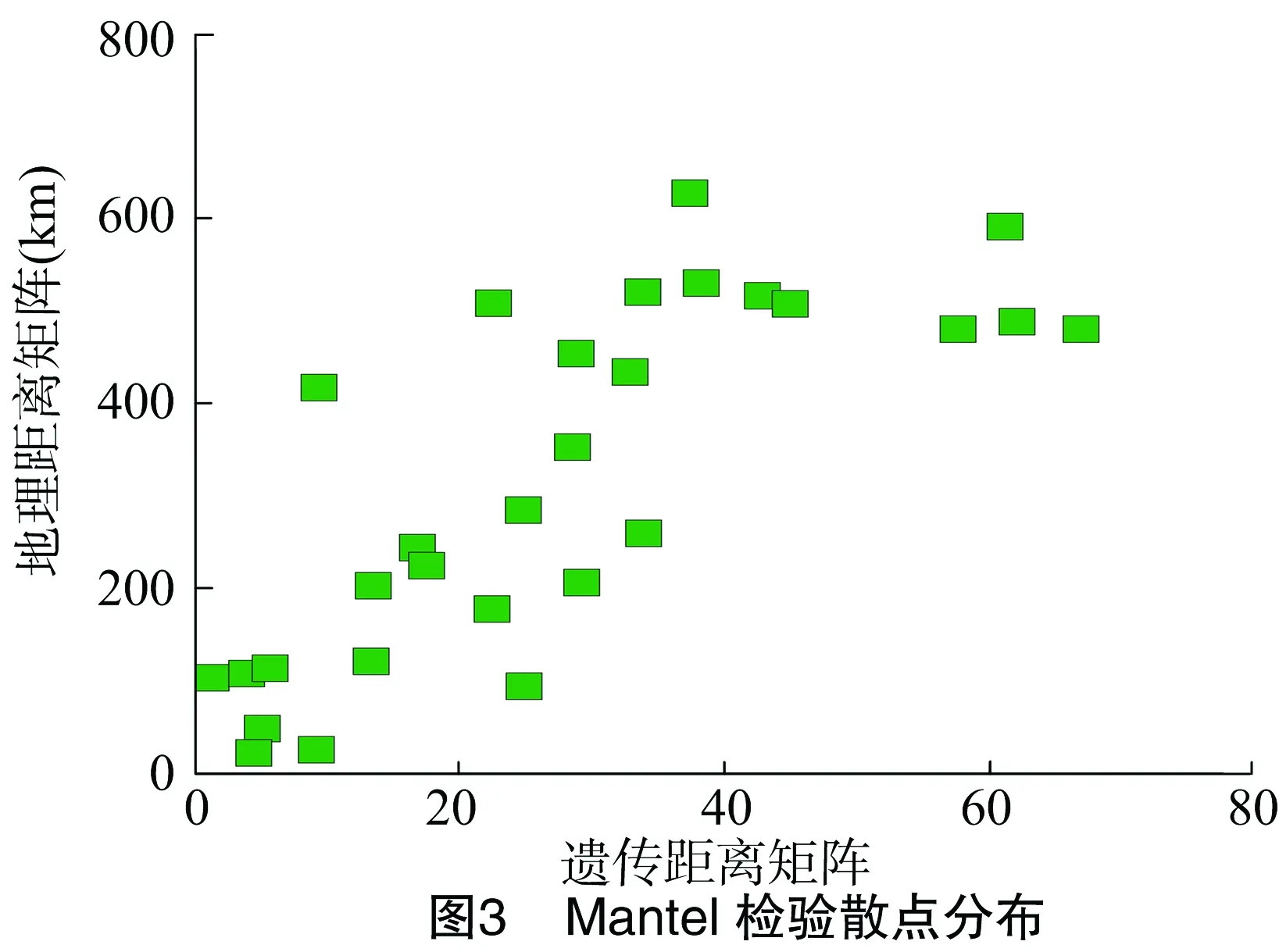
2.5.3 表型性状间遗传距离与地理距离检验 湖北不同种源红椿8个居群种实表型性状的遗传距离与地理距离经过Mantel检测(表8),R=0.783,P=0.002<0.01,说明各居群种实表型变异为显著地理隔离模式,Mantel检测相关关系散点分布见图3。Mantel检测表明,居群间种实表型变异为显著地理隔离模式。
3 结论与讨论
3.1 红椿种实变异丰富性
种实表型性状主要受遗传因素控制,在不同地理分布区也会因适应不同的生境而产生多样性[13]。这主要由于栖息环境条件差异较大,在长期的进化过程中,造成的地理隔离、生境片段化和基因交流不频繁,都会促使一定表型变异的产生[21]。研究发现,不同红椿居群间果纵径、果形指数差异性较大。种实表型性状在种群内的变异系数平均值为3.504%~13.366%,而居群间种实表型性状变异系数为6.400%~31.355%,居群间的变异远大于居群内部。果纵径为最大性状变异(P<0.01)(F=116.646),其次为果形指数(F=106.106)、单果质量(F=74.750)、单种质量(F=82.339),均达到极显著差异(P<0.01)。表型多样性指数表明,通山九宫山、黄石黄荆山、谷城玛瑙观多样性指数整体偏高,地理变异趋势明显。由表型性状与地理环境因子间的相关性分析可知,红椿种子质量受地理空间经度变化影响较大,纬度、海拔变化对种实影响不大,但通过日照时数表现出影响,即随纬度增大、海拔升高,单果质量下降。红椿种实表型性状与地理环境因子之间的相关性分析结果表明,种子质量受年积温效应影响较为明显,积温相对较高的生长环境更适于种子质量的累积,这反映出红椿强阳性的生物学特性。主成分分析和聚类分析结果表明,通山九宫山居群与黄石黄荆山居群遗传距离较近,鄂西北地区的2个居群遗传距离较近,鄂西南地区的4个居群遗传距离较近,与地理方位一致,说明湖北不同种源红椿居群种实表型变异为显著地理隔离模式。

表8 不同居群红椿种实遗传距离与标准化地理距离
注:表中下三角数据为不同居群种实遗传距离;上三角数据为标准化ln(x+1)地理距离。
3.2 红椿遗传育种展望
湖北不同种源红椿种实表型特征的多样性具有地理适应意义,其地理变异特征随经度的变化而变化,与纬度无明显相关性。果纵径、果横径、单果质量、单种质量等主要性状指标明显随经度增高而递减,说明红椿种实的变异与自然分布区内环境异质性有关。湖北为红椿在亚热带自然分布的北缘地带[22],湖北境内环境异质性较大,中部跨越江汉平原,生态气候类型差别较大,不同局部立地环境差异十分明显。鄂西地区红椿居群多处于自然保护区,地理环境、生物多样性更为丰富。鄂中地区为平原和低山丘陵,居群自然分布受到阻隔。鄂东南分布区为低山区和低山丘陵,红椿天然居群表型性状与鄂西地区差异明显。地理隔离造成生态类型的差异,使得红椿不同地理区域居群间种实表型性状自然变异丰富,对天然居群保持较高的遗传多样性是有利的。
表型变异是遗传型和环境因子共同作用的结果,表型变异必然蕴藏着遗传变异。表型变异越大,可能存在的遗传变异越大[23]。地理隔离与自然选择使得湖北不同种源红椿产生了极其丰富多样的地理种源变异,其改良前景很大。因此,在种源苗期试验和造林试验工作中,应该注重优良种源和优良个体选择,这对于湖北地区红椿种质资源的保护、评价和利用具有现实意义。
