浙江省极小种群多脉铁木年龄结构与动态
2018-08-24吴世斌库伟鹏周小荣周海伟纪美芬吴家森
吴世斌,库伟鹏,周小荣,周海伟,纪美芬,吴家森*
(1.文成县林业局,浙江 文成 325300; 2.浙江农林大学 环境与资源学院,浙江 杭州 311300; 3.文成县苗圃,浙江 文成 325300; 4.文成县石垟林场,浙江 文成 325300; 5.文成县叶胜林场,浙江 文成 325300)
植物种群结构是指种群内不同年龄、大小个体的分布状况,反映了各龄级个体的组配、立地条件和环境间的相互关系[1]。生命表和存活曲线能直观地展现不同龄级个体的现有的生存数、死亡数[2-3],分析过去的种群结构和受干扰情况,预测未来的种群变化,了解种群对环境条件的适应性[4],对于濒危植物的保护和管理具有重要意义。
多脉铁木(Ostryamultinervis)为桦木科铁木属植物,分布于湖北利川、湖南紫云山、四川东南部和贵州等地,其分布区孤立局限,具有树体高大,干形通直,材质优良的特点[5],为我国特有种植物,是浙江省珍稀濒危野生植物和极小种群保护对象。已有研究对多脉铁木的繁育技术、林分特征进行了初步研究[6-7],而有关多脉铁木种群的年龄结构及数量动态的研究未见报道。本文详细调查了浙江省文成县石垟林场多脉铁木群落的所有个体,利用径级结构分析,编制了种属静态生命表和种群存活曲线,预测种群时间序列的变化趋势,旨在为多脉铁木的种群恢复策略的制定提供理论和参考价值。
1 研究地区与方法
1.1 研究区概况
研究区位于浙江省文成县石垟林场猴王谷景区倒臼源,多脉铁木的分布中心地理位置为27°50′N, 119°50′E。研究地所在的石垟林场属中亚热带季风性气候,年平均气温12.8 ℃,最冷月(1月)平均气温为4.0 ℃,极端最低气温-14.0 ℃,最热月(7月)平均气温为23.6 ℃,极端最高气温37.3 ℃,年降水量1 604 mm,年均相对湿度84.0%,年日照时数1 755 h,全年无霜期288 d[8]。该区域属天然常绿阔叶林,森林覆盖率达90%以上。多脉铁木群落分布区域的海拔介于500~700 m,坡度30°~35°,南坡,坡中部,地表有石块层覆盖,厚度20~30 cm,石块大小20~40 cm。土壤为发育于花岗岩的红壤土类,表层土壤(0~20 cm)理化性质为pH值4.6,有机碳63.2 g·kg-1,碱解氮、有效磷、速效钾含量分别为145.2、21.82、187 mg·kg-1。
多脉铁木种群中植物资源丰富,共有维管植物60种(含变种),隶属于40科52属。伴生的乔木树种有8种,分别是褐叶青冈、小叶栎、四照花、缺萼枫香、短尾柯、钩栲、尾叶冬青、麂角杜鹃;灌木层主要植物有披针叶茴香、毛花连蕊茶、光叶石楠、长裂葛萝槭、红楠、三尖杉、猴欢喜、大叶冬青、秀丽槭、青榨槭;草本层植物主要有里白、黑足鳞毛蕨、麦冬、求米草、三穗薹草、瘤足蕨、江南卷柏、毛叶腹水草、大叶唐松草、延羽卵果蕨。
1.2 研究方法
1.2.1 调查
2017年8月上旬,全面调查种群内所有的多脉铁木个体,对其所有个体进行每株检尺(起测径阶≥5.0 cm),测定树高、胸径、冠幅,记录<5.0 cm的幼苗数量,并访问林场老职工,了解该种群分布历史及利用情况。
1.2.2 径阶结构划分
根据多脉铁木的生物学特性和野外调查数据的特点,将多脉铁木种群划分为10个径阶,每个径阶对应相应的龄级。径阶<5 cm为第Ⅰ龄级,5~<10 cm为第Ⅱ龄级,10~<15 cm为第Ⅲ龄级,15~<20 cm为第Ⅳ龄级,20~<25 cm为第Ⅴ龄级,25~<30 cm为第Ⅵ龄级,30~<35 cm为第Ⅶ龄级,35~< 40 cm为第Ⅷ龄级,40~<45 cm为第Ⅸ龄级,D≥45 cm为第Ⅹ龄级。
1.2.3 数据分析
多脉铁木种群的年龄结构、静态生命表及存活曲线、动态量值及指数、时间序列预测等指标参考申仕康等[1]的方法进行。
2 结果与分析
2.1 多脉铁木种群的年龄结构
调查研究表明,多脉铁木种群在其全分布范围内共找到118株,其年龄结构如图1所示。从图中可知,Ⅰ龄级的幼苗有13株,占总数的11.0%,而Ⅱ龄级的幼树仅1株,而后随着年龄的增大,植株个体数量先增加而后下降,多数集中于Ⅴ~Ⅶ龄级,占整个种群的52.%。多脉铁木的年龄结构近似正态分布的特点,种群暂时稳定。但Ⅱ龄级个体数量急骤减少,表明该种群在演替过程中受到严酷的环境筛影响,导致Ⅰ龄级向Ⅱ龄级过渡中死亡率达92.3%,严重影响着种群的发展。

图1 多脉铁木的龄级结构
2.2 多脉铁木种群的静态生命表及存活曲线
从静态生命表(表1)和存活曲线(图2)中可知,多脉铁木幼苗个体数量较多,种群存活曲线趋于Deevey-Ⅲ型,多脉铁木个体由第Ⅰ龄级向第Ⅱ龄级演替转化的过程中,其数量显著下降,仅有7.7%的个体能进入到第Ⅱ龄级生长,这表明幼年个体向成年阶段的发育是不连续的,该时间已成为多脉铁木种群发展的“瓶颈”。多脉铁木Ⅲ~Ⅶ龄级的个体数目先增加而后保持稳定,到第Ⅷ龄级后,其个体数量又呈现减少趋势。

表1 多脉铁木种群的静态生命

图2 多脉铁木种群存活曲线

图3 多脉铁木种群死亡率(Qx)和损失度(Kx)曲线
多脉铁木种群死亡率(Qx)和损失度(Kx)的变化趋势相似(图3),Kx在第Ⅰ龄级和第Ⅷ龄级出现峰值,第Ⅰ龄级的峰值是因为幼苗以高损失率为代价向第Ⅱ龄级转换,这与存活曲线的变化规律是一致的;第Ⅷ龄级的峰值是由于存活个体的生理衰老退化,从而形成损失高峰。多脉铁木Ⅲ~Ⅶ龄级的Qx和Kx接近于0,表明该年龄段其种群结构稳定。
2.3 多脉铁木种群的动态量值

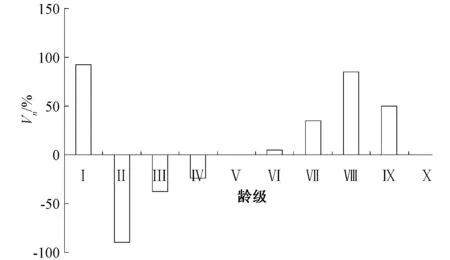
图4 多脉铁木种群的动态量值

图5 多脉铁木种群动态指数
2.4 多脉铁木种群的时间序列预测
多脉铁木的时间序列预测(表2)表明,从2019年起,在之后的2 a内,多脉铁木种群Ⅱ龄级以上的个体数量呈现增长趋势,而5 a后,该物种仅在Ⅴ龄级后才有个体增加,表现为低龄级的个体数量不足,种群面临衰退。

表2 多脉铁木龄级结构的时间序列预测
3 小结与讨论
某一植物种群的年龄结构是其生物学特性与环境因子共同作用的结果[9],年龄金字塔呈纺锤形的种群为稳定的群落结构[10]。本研究结果表明,多脉铁木种群从第Ⅱ龄级开始的年龄结构金字塔基本呈纺锤形,近似于正态分布,即中龄个体数量占种群总体数量的比例较高,而幼龄和老龄个体较少,表现为暂时的稳定结构特征。尽管Ⅰ龄级的多脉铁木幼苗数量较多,但也仅13株,而Ⅱ龄级的幼树数量仅1株,出现明显的瓶颈现象,受到环境筛的显著影响。种子产量低、品质差,种子向幼苗的转化率低,形成的幼苗数量少是濒危植物存在的较普遍问题[11]。多脉铁木种子的种皮坚硬、透水性差,吸水主要通过苞片网脉,种子不育、空瘪粒多,种子饱满率仅12%。因此种子品质差是造成多脉铁木幼苗少的内在因素;其次,多脉铁木种群所在的立地环境中,表层有大量石块覆盖,种子较轻、有种翅,成熟后随风散布,很难进入土壤层,种子难以在土壤上着床,不能萌芽成苗,也是造成多脉铁木幼苗较少的关键外在因素。另外随着生态公益林保护力度的增大,多脉铁木群落各种树木的生长,森林的郁闭度达90%以上,喜光的多脉铁木幼苗得不到足够的光照条件,因而在Ⅰ、Ⅱ龄级遭遇1次死亡高峰。从第Ⅲ龄级开始,多脉铁木种群保持较高的稳定性。
种群生命表可以判断植物种群发展趋势,存活曲线能够直观反映种群个体在各龄级的存活状况[12],Deevey-Ⅲ型存活曲线主要表现为早期死亡率高,一旦存活到某一年龄,死亡率就较低,可视为增长种群[13]。本研究结果表明,多脉铁木种群的存活曲线属于Ⅲ型,Ⅰ、Ⅱ龄级的幼龄个体受到严酷的环境筛作用,对幼龄树的生存不利;但到Ⅲ龄级后,种群呈增长型结构,这与种群的动态量值和动态指数的结果也是一致的。从种群的时间序列分析,5 a后,该物种在Ⅴ龄级前均没有新的个体增加,低龄级的个体数量严重不足,种群可能面临衰退。
针对多脉铁木种群的现状和发展趋势,可从以下两方面考虑该种群的保护和恢复策略。一方面从种群个体自身因素考虑,在加强对原生境及植株的保护与监测的基础上,加大科研力度,开展多脉铁木生殖生态学研究,如种子萌发率、成苗率和高效繁育技术等,提高实生苗的数量,以扩大其种群分布范围;另一方面是采取“近地保护”措施,将人工繁育的实生苗在多脉铁木现有分布区附近进行适当种植,建立人工保护点。从种群所处生境考虑,适度开展人为干预,搬动部分大石块,展露部分区域土壤,以保证多脉铁木种子顺利入土,促进种子萌发。在此基础上,间伐过密的非目标树种,适当扩大林窗,以降低森林郁闭度,增加林内光照,以利于幼苗的生长,从而突破多脉铁木由Ⅰ龄级向Ⅱ龄级转化的“瓶颈”。
