一种由山葵果胶杆菌引起的马铃薯细菌性软腐病
2018-08-16王亚艺高旭升蔡晓剑
王 信,程 亮,王亚艺,高旭升,蔡晓剑
(青海大学农林科学院,青海 西宁 810016)
【研究意义】马铃薯是我国第四大粮食作物,也是重要的蔬菜和加工原料。2013年国家实施马铃薯主食化战略后[1],主产区省(自治区、直辖市)对马铃薯生产和管理更加重视,马铃薯种植面积稳步扩大,近年来基本稳定在550万hm2左右。南方主要将其作为蔬菜,北方则作为主要的粮食作物,尤其在西北干旱、半干旱地区及西南高寒山区马铃薯成为了主要的高产和救灾作物。马铃薯是青海省主要的粮食作物之一,2013年青海省马铃薯种植面积达到9.56万hm2,总产量达到2150万t[2],仅次于小麦和油料,成为第3大作物,青海的马铃薯主要种植于互助、民和、乐都、湟中、大通、平安等东部各县的山区。其中马铃薯软腐病是青海马铃薯的主要病害之一,在田间、运输途中及贮藏期间均可发生软腐病,并常与干腐病复合侵染,引起较大损失,损失率达3 %~68 %,平均为15 %以上[3-4],随着带病种薯的频繁调运,种植面积逐年扩大,该病的危害越来越重。【前人研究进展】马铃薯软腐病是由几种果胶杆菌属单独或复合侵染所引起的,通常情况下会对马铃薯块茎造成严重危害。不同国家和地区对引起马铃薯腐烂的病原菌的相关报道不尽相同[5-6]。目前,公认的引起马铃薯软腐病的病原主要是由Dickeyaspp.[7-8]和Pectobacteriumspp.[5-6]。Dickeya是近年来成立的植物病原细菌新属,其模式菌为原来的菊欧氏菌(Erwiniachrysanthemi)。Dickeyaspp.引起的细菌性软腐病常给作物和花卉产业带来严重的经济损失。该菌所引起的马铃薯细菌性软腐病近年造成了欧洲国家、以色列和美国的乔治亚州马铃薯的大幅减产[7-8],在我国马铃薯产区也有发生,该病是一种新发现的随马铃薯种薯传播的细菌性病害,比传统的黑胫病菌(E.carotovorumsubsp.atrosepticum)侵染性更强,对欧洲的马铃薯生产已构成威胁[7-8]。果胶杆菌属(Pectobacteriumspp.)依据其寄主范围、生理生化和分子特征,将其分为5个种,分别为Pectobacteriumcarotovorumsubsp.carotovorum(Pcc) (syn.Erwiniacarotovorumsubsp.carotovorum),P.atrosepticum(Pa) (syn.E.carotovorumsubsp.atrosepticum),P.wasabiae(Pw) (syn.E.carotovorumsubsp.wasabiae),P.betavasculorum(Pbv) (syn.E.carotovorumsubsp.betavasculorum)和P.carotovorumsubsp.brasiliense(Pcb)[9]。据相关文献报道,它们的地理分布具有很大差异。其中Pcc作为引起马铃薯软腐病的主要病原菌,其次是Pa、Dickeyaspp则较少。在我国主要是以Pcc和Dickeyaspp为主,Pa较少[10]。80年代中期,王金生等也曾报道马铃薯软腐欧文氏杆菌Pa、Pcc和Dickeyaspp在我国的组成及分布,在284个菌株中Pcc占72.5 %,为优势病原菌;而集中分布在我国北方地区的Pa病原菌仅占5.3 %[11]。其中Pw寄主范围很窄,最早在日本的辣根上分离出来,后来在美国、新西兰和伊朗的马铃薯和观赏植物中得到[12-16]。然而,在我国马铃薯和其它观赏性植物上分离到Pw文献鲜有报道。【本研究切入点】本研究对大通县朔北乡多隆村马铃薯生产地块马铃薯软腐病发病组织中分离获得菌株QDD-5进行致病性分析,发现QDD-5在离体和活体条件下均能引起马铃薯块茎组织典型的软腐症状,经表型特征及16S rRNA基因序列系统分析,该菌株被鉴定为Pw。这是首列引发青海马铃薯软腐病病原菌报道,该病原菌鉴定对于控制马铃薯软腐病,降低果胶杆菌对马铃薯产业的影响具有重要意义。【拟解决的关键问题】明确青海省马铃薯细菌性软腐病的系统发育和分布特点。
1 材料与方法
1.1 病样采集及培养基
马铃薯软腐病标样采集于青海省的大通县的马铃薯生产田。 结晶紫果胶酸盐选择性培养基(CVP):在500 mL的沸腾的蒸馏水中按顺序加入:1 mL 0.75 g/L 结晶紫溶液,4.5 mL 1 mol/L NaOH溶液,3 mL 100 g/L CaCl2·2H2O溶液(新鲜溶液),3.0 g琼脂粉,1.0 g NaNO3,9.0 g聚果胶酸钠,待聚果胶酸钠加入时人工搅拌30 s,然后分别装入三角瓶,121 ℃灭菌20 min,趁热倒将培养基入无菌培养皿中,冷却后制成平板。
Luria-Bertani培养基(LB):胰蛋白胨10.0 g,酵母提取物5.0 g,氯化钠10.0 g、琼脂粉15 g,用去离子水混合定容至1000 mL,pH 7.0。然后分别装入三角瓶,121 ℃灭菌20 min,趁热将培养基倒入无菌培养皿,冷却后制成平板。
1.2 病原菌分离
采用常规组织分离法分离病原菌。称取0.1 g病块茎置于250 mL的三角瓶中,并加入100 mL的蒸馏水,放入玻璃珠充分打散混合液,置于120 r/min 摇床中,培养30 min后,充分摇匀。用1 mL无菌的吸管吸取10-2菌悬液1 mL放入9 mL的无菌水管中,吹吸3次混匀即为10-3稀释液,依次从10-3连续稀释至10-7,即得一系列的10倍的稀释液。每个梯度重复进行3次平行试验,分别涂布到CVP培养基上,25~27 ℃黑暗培养48 h,将典型形态的菌落在CVP培养基反复划线,进行纯化培养,挑取单菌落进行编号和保存。
1.3 致病力测定
选择健康马铃薯品种大西洋的块茎用于致病性分析。将待测菌株于液体LB培养基中培养24 h,用无菌的9 g/L NaCl溶液(w/v)洗涤3次后,配置5×108cfu/mL的菌悬液。将无病的大小均匀的块茎用自来水冲洗后放入0.5 %NaClO溶液(v/v)中洗2次,每次20 min,然后用无菌去离子水冲洗1~2次,放入超净台中自然晾干,用无菌竹牙签给每个块茎打2~3个接种孔,深度约为20 mm,取10 μl菌悬液用无菌9 g/L NaCl溶液配制成5×108cfu/mL的菌悬液加入到接种孔中,每个株系接种10个块茎。用凡士林将块茎上的接种孔封口保湿,将接种的块茎置于温度为22 ℃、相对湿度≥92 %的温室培养。同等条件下,设无菌9g/L NaCl溶液为空白对照。接种后72 h时测量病斑的直径并取其平均值作为病斑平均直径。按以下分级标准记录病斑:小于1~5 mm为弱致病力,5~10 mm为中等致病力,10 mm以上为强致病力。
1.4 株系形态和生物化学特性分析
菌落形态、革兰氏染色和鞭毛染色及柠檬酸盐分解和37 ℃生长测定参考《植病研究方法》(第3版)[17]进行。病原菌对蔗糖、麦芽糖、山梨醇、阿拉伯醇、乳糖和α-甲基葡糖苷等的利用测定实验参考《伯杰氏细菌鉴定手册》(第8版)[18]和《植病研究方法》(第3版)[17]进行。选用本实验室保存的软腐病致病菌Pcc(编号: ECC71)为对照株系。
1.5 16S rRNA基因片段的扩增和序列分析
1.5.1 基因组DNA的提取 将保存于-80 ℃冰箱的株系划线并于28 ℃培养箱中倒置培养过夜活化,挑取单菌落放入液体LB培养基中置28 ℃培养16~18 h。病菌DNA的提取使用Ezup柱式SK8255试剂盒(生工生物工程(上海)股份有限公司)。取1 μl DNA溶液于2.0 %琼脂糖凝胶(w/v)进行电泳检测,并于-20 ℃保存,备用。
1.5.2 16S rRNA片段的扩增 应用细菌通用引物27f(5′-AGAGTTTGATCGGCTCAG-3′;1942 r(5′-TACGGCTACCTTGTTACGACTT-3′)进行扩增。每个PCR反应体系总体积均为25 μl,包括2×TaqPCR MasterMix 12.5 μl(编号为KT201-02,Tiangen),DNA模板(50 ng/L)1 μl,引物 27f 和引物1942 r (μmol/L) 1 μl,加双蒸水10.5 μl。PCR扩增程序:95 ℃变性5 min;95 ℃ 30 s,52 ℃ 30 s,72 ℃ 1.5 min,40个循环;72 ℃延伸10 min。回收大小约1.5 kb的PCR产物。将其连接到pGEMT-Easy载体(Promega corp.,美国)上,转化到大肠杆菌(Escherichiacoli)DH5α感受态细胞中。重组质粒经NotⅠ酶切和PCR鉴定后,送交生工生物工程(上海)股份有限公司测序。
1.5.3 16S rRNA系统发育树构建 测序结果用软件Chromas(ver. 2.4.1)进行序列修正后,在NCBI数据库进行Blast检索与目标序列同源性高的DNA序列,与本文所测株系序列分别进行序列比对。利用软件MEGA(ver. 5.05)的UPGMA法构建系统发育树,各分支的置信度自举检测(bootstrap)1000次。
1.6 Pw特异引物扩增基因
Pw特异性引物EXPCCF(5′-GAACTTCGCACCGCCGACCTTCTA-3′)和EXPCCR (5′-GCCGTAATTGCCTACCTGCTTAAG-3′)[19]用于扩增2个分离株系pECC2F序列区域。25 μl的PCR反应体系:DAN模板1 μl(100 ng/L),2×TaqPCR Master Mix 12 μl,上游引物1 μl(20 μmol/L),下游引物1 μl(20 μmol/L),加ddH2O至总体积25 μl。PCR扩增条件: 94 ℃变性3 min;94 ℃30 s,56 ℃ 30 s,72 ℃ 1.5 min,30个循环;72 ℃延伸5 min。以2000 bp DNA Marker为分子质量标准,取4 μl PCR扩增产物进行2.0 %琼脂凝胶电泳(w/v),核酸染料染色观察是否有特异性条带产生。选用本实验室保存的果胶杆菌致病菌Pcc(ECC71)为对照株系。
2 结果与分析
2.1 Pw的分离和致病性测定
QDD-5病样分离的菌株经CVP选择性培养基筛选,在CVP培养基上生长的株系向培养基内部扩展,形成杯状凹陷,将分离的株系回接到宿主马铃薯块茎上,块茎组织从接种部位开始产生水渍状病变并向接种点周围迅速扩散(图1-B)。对接种发病的马铃薯块茎组织重新分离病原菌,得到的1株能引起马铃薯块茎软腐症状。
2.2 软腐病菌的形态及生物化学特征分析
软腐病病样中分离的QDD-5经CVP选择性培养基筛选,在CVP培养基上产生凹陷的株系。将株系转接到LB固体培养基上,菌落形态呈圆形、乳白色、表面光滑,略隆起(图1-A)。革兰氏染色呈阴性。测试菌株QDD-5不能在37 ℃ 生长,能使明胶液化,不具有耐盐性,对红霉素不敏感,过氧化氢酶反应为阳性,氧化酶、蔗糖试验还原反应、吲哚产生和磷酸酶活性试验均为阴性,可以利用纤维二糖和柠檬酸盐,不能利用D-麦芽糖、山梨醇、D-阿拉伯糖、α-甲基葡萄糖、乳糖、棉子糖和蜜二糖(表1)。这些结果与报道的P.wasabiae(黑腐果胶杆菌)的生物学形态特征和生理生化特性均吻合[15]。
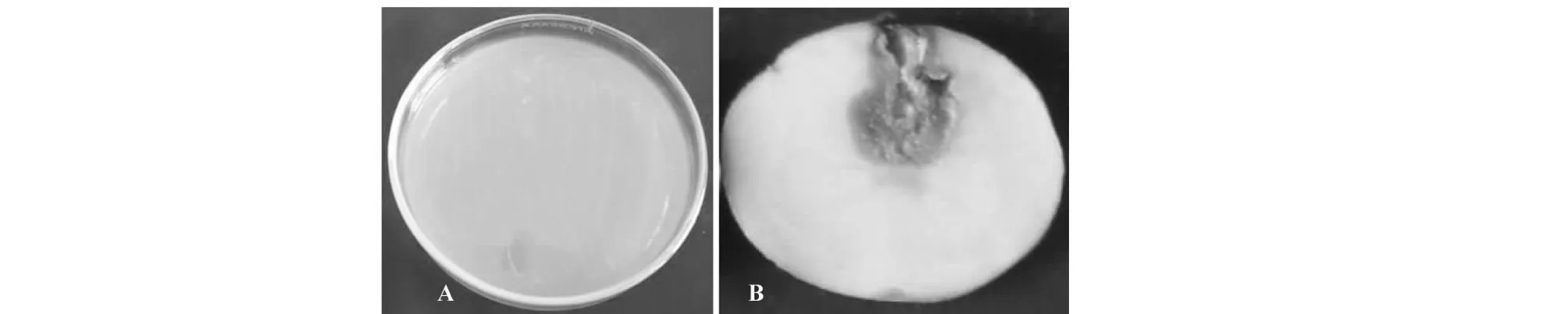
图1 株系在LB培养基平板上产生乳白色、光滑隆起菌落(A)和接种72 h后块茎症状(B)Fig.1 Cup-shaped cavities formation of isolated pathogen from diseased tuber of potato in field on LB agar (A) and symptom of healthy tuber at 72 hours after in vitro-inoculation with the pathogen (B)

表1 菌株QDD-5的生理生化特性
注:+,阳性反应;-,阴性反应。*,马铃薯软腐病致病菌株Pcc,Pectobacteriumcarotovorumsubsp.carotovorum(ECC71),由本实验室保存并提供本研究中作为阳性对照使用。
Note: ‘+’and ‘-’ indicated positive and negative reactions, respectively. *:One known strain causing bacterial soft rot of potato,Pcc,Pectobacteriumcarotovorumsubsp.carotovorum(ECC71), were preserved in our laboratory and provided to use as positive control in this study.
2.3 16S rRNA的序列分析及系统发育树构建
本研究中分离获得的QDD-5和对照菌株Ecc71的16S rRNA基因PCR扩增,均得到一条大小约为1.5 kb的产物(图2),测序结果经在NCBI数据库中与已知序列进行BLAST比对,序列比对结果表明,菌株QDD-5与已报道的Pw(NR116048.1)的16SrRNA基因的序列的相似度(或一致性)均达99 %。基于16S rRNA基因完整序列,以马铃薯黑胫病致病菌DickeyadadantiiEch586(CP001836.1)为外组,将QDD-5的序列与已发表的28个株系的序列构建系统发育树(图3)。结果显示,本研究分离得到的QDD-5与已发表的Pw株系形成的类群构成了明显的分枝、已发表的Pcc、Pcb与Pco株系形成的类群构成了独立的分枝,Pa和Pb株系共同组成的类群与Pw株系组成的类群也聚集成另外分枝。
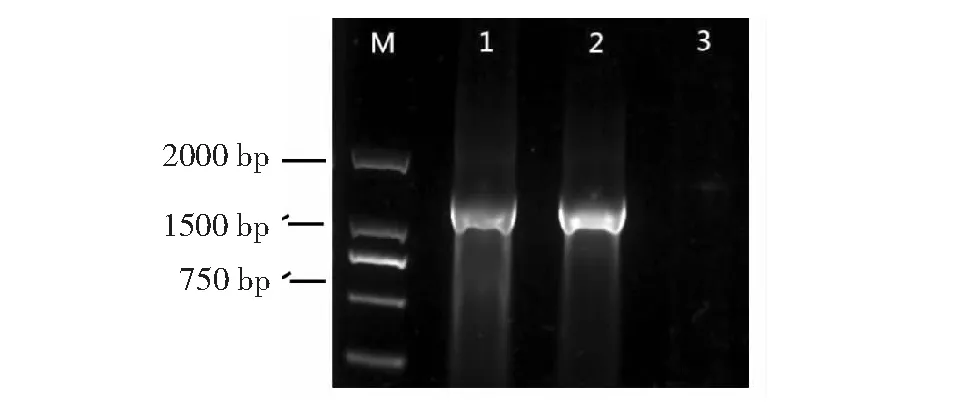
M:DNA marker DL2000;1:QDD-5;2:ECC71;3:无菌水用作阴性对照图2 用引物27f/1942r进行PCR扩增不同分离株系16SrRNA的琼脂糖凝胶电泳Fig.2 Ethidium bromide-stained agarose gel electrophoresis showing PCR products amplified with primers 27f/1942r in 16SrRNA genes of 2 different strains
2.4 Pw特异性引物扩增
用特异性引物EXPCCF和EXPCCR扩增QDD-5及其对照菌株Ecc71。结果表明这2个菌株中均扩增得到一条大小约为550 bp的特异条带(图4);由此可见,特异性引物EXPCCF和EXPCCR既能扩增Pcc,又是Pw的特异性引物。
2.5 软腐病菌致病力测定
本研究以马铃薯块茎接种病原菌产生的病斑长度作为细菌致病力的测定,块茎竹签伤口接种后72 h统计病斑长度,发现QDD-5与Ecc71之间不存在明显的致病力差异,按病斑长度划分低、中和高等致病力。在接种后72 h时表现较强致病力,QDD-5与Ecc71致病力均是中等(表2)。
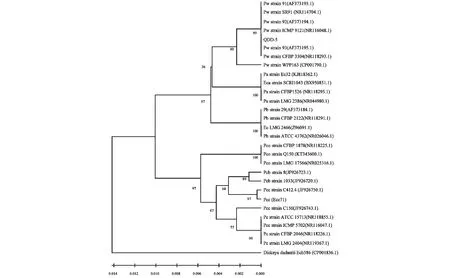
每个分支点的数目显示自举支持的百分比。bar表示进化枝的遗传距离图3 菌株QDD-5与其他株系的16S rRNA基于序列系统发育树Fig.3 Phylogenetic tree based on comparison of complete 16S rRNA gene sequences of QDD-5 strain with that of 28 Pectobacterium strains submitted at NCBI database
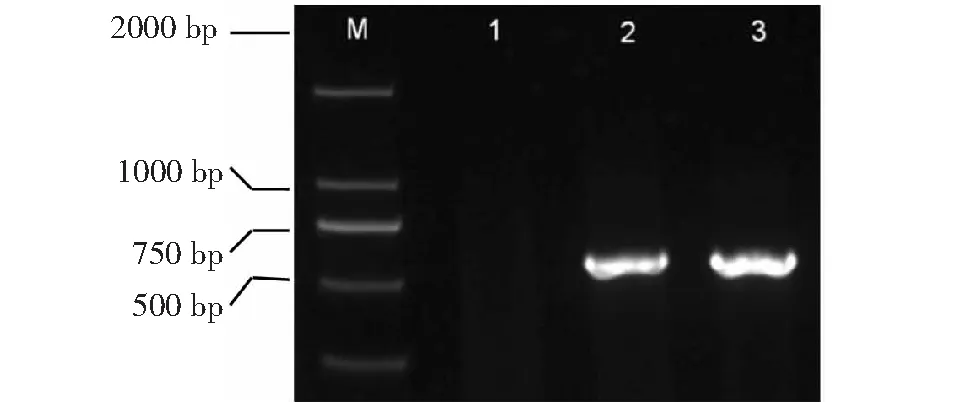
M:DNA marker DL2000;1:无菌水用作阴性对照;2:QDD-5;3:ECC71图4 分离的2个用特异引物EXPCCF/EXPCCR的PCR扩增产物琼脂糖凝胶电泳Fig.4 Ethidium bromide-stained agarose gel electrophoresis of PCR products amplified with primers EXPCCF/EXPCCR from 2 different strain collections
3 讨 论
本研究对从大通县朔北乡多隆村马铃薯软腐病样中分离到的菌株QDD-5进行了形态和生理生化表型特征观察以及分子诊断,将其鉴定为Pectobacteriumwasabiae。Pw的发生引起了包括欧洲、亚洲、澳洲、非洲和美洲国家的密切关注。据调查,Pw在20世纪70年代波兰和荷兰马铃薯植株上发生,80年代在美国分离获得PwIFB5311、IFB5312和IFB5313菌株,90年代初在芬兰和苏格兰确认该病原菌。本试验菌株QDD-5与对照菌株ECC71生理生化指标存在着许多差别,最明显的区别是测试菌株QDD-5不能在37 ℃生长,不具有耐盐性,不能利用乳糖、棉子糖和蜜二糖,而ECC71则刚好与其相反,所以这2个菌株也属于不同的亚类群,QDD-5与发表的Pw株系表现一致,支持其属于Pw种的归类。
随着分子生物技术的发展,16S rRNA已经发展成为用于表型特征无法解决的Pectobacterium种或亚种的鉴别。为了准确特征分离到的QDD-5菌株,本研究克隆了QDD-5和ECC71菌株16S rRNA基因序列,并将其与发表的27个软腐果胶杆菌16S rRNA基因序列类型比对分析,发现QDD-5与已发表的6个株系相似性达到99 %,共同形成明显的Pw分枝。Edward 等报道用EXPCCF/EXPCCR特异性引物在Pcc菌系中扩增到550 bp的特异性条带(ECC71),而Pw中有相同的扩增产物[19](QDD-5)。Nabhan等研究认为16SrRNA仅能区分细菌种的差异,而不能对亚种进行区分[20],本研究中在QDD-5和ECC71经过16S rRNA完整序列扩增和UPGMA法构建系统发育树分析,聚成不同类群,QDD-5菌株和Pw株系ICMP 9121(NR116048.1)构成亚类群,而ECC71与Pcc(JF926743.1)聚在一起构成亚类群,遗传关系相对较远的Pco和Pcb株系单独成群。可见,基于16S rRNA基于序列在一定程度上也揭示亚种之间的遗传关系。在以前研究中,马铃薯上的Pw分离物能使马铃薯薯块软腐、茎枯萎或褐化[14,21-22]。然而,用PCR方法或DNA杂交的结果显示[13],Pw缺失三级分泌系统(T3SS)或者缺失T3SS基因簇的调节基因,而T3SS是革兰氏阴性细菌中重要的在不同环境中传递毒性因子的系统,也是Pa、Pc等果胶杆菌侵染马铃薯参与致病性的基因簇[23-24]。然而,T3SS对Pw的致病性或许没有像在Pa、Pc中那么重要,或者Pw是用其它基因簇代替T3SS侵染寄主植物。Pw除了T3SS缺失,其它分泌系统是否和果胶杆菌其它亚种相同,还需要更进一步研究。同时从马铃薯软腐性病害分离到的主要是Pcc,黑胫病分离到的主要是Pa,关于Pw、Pcc与Pa二者或三者在青海马铃薯产区的分布、扩散及复合浸染也有待进一步研究。本研究对青海马铃薯细菌性软腐病病原进行了准确鉴定,也是首次报道由Pw引起青海马铃薯软腐病,这一研究结果可为马铃薯软腐病综合控制技术研究奠定基础。

表2 Pw株系的采集地点及致病力等级
注:M为致病力中等。
Note: M represents moderate pathogenicity.
4 结 论
综上所述,Pectobacterium是引起青海地区马铃薯软腐病的主要病原菌,在马铃薯生长、收获、运输和贮藏期间,都可以造成重大损失。从青海省大通县马铃薯软腐组织中分离到软腐病原菌1株。基于生物学鉴定和16S rRNA序列以及特征序列扩增分析,该菌株在系统发育上属于Pectobacteriumwasabiae。生理生化试验表明,该病原菌株不能在37 ℃ 生长,能使明胶液化,不具有耐盐性,对红霉素不敏感,过氧化氢酶反应为阳性,氧化酶、蔗糖试验还原反应、吲哚产生和磷酸酶活性试验均为阴性,可以利用纤维二糖和柠檬酸盐,不能利用D-麦芽糖、山梨醇、D-阿拉伯糖、α-甲基葡萄糖、乳糖、棉子糖和蜜二糖。
