Preliminarily study on the maximum handling size, prey size and species selectivity of growth hormone transgenic and non-transgenic common carp Cyprinus carpio when foraging on gastropods*
2018-08-02ZHUTingbing朱挺兵ZHANGLihong张丽红ZHANGTanglin张堂林WANGYaping汪亚平HUWei胡炜OLSENRolfEricZHUZuoyan朱作言
ZHU Tingbing (朱挺兵) , ZHANG Lihong (张丽红) ZHANG Tanglin(张堂林) ,WANG Yaping (汪亚平) HU Wei (胡炜) OLSEN Rolf Eric , ZHU Zuoyan (朱作言)
1 State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences,Wuhan 430072, China
2 Yangtze River Fisheries Research Institute, Chinese Academy of Fisheries Science, Wuhan 430223, China
3 Department of Biology, Norwegian University of Science and Technology, 7491 Trondheim, Norway
Abstract The present study preliminarily examined the differences in maximum handling size, prey size and species selectivity of growth hormone transgenic and non-transgenic common carp Cyprinus carpio when foraging on four gastropods species ( Bellamya aeruginosa, Radix auricularia, Parafossarulus sinensis and Alocinma longicornis) under laboratory conditions. In the maximum handling size trial, five fish from each age group (1-year-old and 2-year-old) and each genotype (transgenic and non-transgenic)of common carp were individually allowed to feed on B. aeruginosa with wide shell height range. The results showed that maximum handling size increased linearly with fish length, and there was no signi ficant difference in maximum handling size between the two genotypes. In the size selection trial, three pairs of 2-year-old transgenic and non-transgenic carp were individually allowed to feed on three size groups of B. aeruginosa. The results show that the two genotypes of C. carpio favored the small-sized group over the large-sized group. In the species selection trial, three pairs of 2-year-old transgenic and non-transgenic carp were individually allowed to feed on thin-shelled B. aeruginosa and thick-shelled R. auricularia,and five pairs of 2-year-old transgenic and non-transgenic carp were individually allowed to feed on two gastropods species ( P. sinensis and A. longicornis) with similar size and shell strength. The results showed that both genotypes preferred thin-shelled Radix auricularia rather than thick-shelled B. aeruginosa, but there were no signi ficant difference in selectivity between the two genotypes when fed on P. sinensis and A. longicornis. The present study indicates that transgenic and non-transgenic C. carpio show similar selectivity of predation on the size- and species-limited gastropods. While this information may be useful for assessing the environmental risk of transgenic carp, it does not necessarily demonstrate that transgenic common carp might have lesser environmental impacts than non-transgenic carp.
Keyword: transgenic fish; Cyprinus carpio L.; fast growth; predation ability; prey selectivity
1 INTRODUCTION
Using transgenic technology to produce targeted genetically modi fied organisms (GMO) is an interesting method to meet the food requirement of a growing world population. Transgenic techniques for enhancing growth have been used since the 1980’s and include fish like Atlantic salmon, Salmo salar and Nile tilapia, Oreochromis niloticus (Maclean and Laight, 2000). China has had a major role in the development of fast growing common carp Cyprinus carpio. One of the most promising advances was the transfer of a genomic growth hormone (GH) construct from grass carp, Ctenopharyngodon idella to C. carpio (Wang et al., 2001). However, very few transgenic fish have been permitted for commercial production. One central concern are the potentially negative environmental effects upon escape or release(Maclean and Laight, 2000). The possible ecological risks of transgenic fish are in many aspects, such as competition with wild species, changes in dietary selection and interbreeding creating new genetic variants (Hu et al., 2007).
The transgenic C. carpio is known to have higher appetite (Duan et al., 2009) and food intake (Fu et al.,2007) to support faster growth. Furthermore, fast growing fish may change their food selection or priorities. Transgenic coho salmon, Oncorhynchus kisutch is known to have both enhanced feeding motivation and increased interactions with novel prey(Sundström et al., 2004). If normal and transgenic C. carpio have different feeding motivation and ability, their prey selectivity may also be affected.
Cyprinus carpio exhibits clear prey selectivity in all growth stages. Fry C. carpio prefer cladocerans(Chakrabarti and Jana, 1990; Lechelt and Bajer,2016), juveniles are avid consumers of meiobenthic organisms (nematodes, micrucrustaceans,oligochaetes) (Weber and Traunspurger, 2014, 2015).While adults C. carpio are omnivorous (Vilizzi et al.,2015) and feed on a wide range of food sources such as macrophytes (Weber and Brown, 2015) or macroinvertebrates including gastropods (Sin, 2006).Large C. carpio tend to choose large Dreissena polymorpha, compared to small C. carpio (Tucker et al., 1996).
Shell morphology and shell strength are important factors that may in fluence prey selectivity in molluscivorous fish. When allowed to choose between Pyrgula annulata, Valvata piscinalis and D. polymorpha,common carp prefer V. piscinalis which has an easier crushed shell, and a higher percentage of organic content (Stein et al., 1975). Crushing of gastropod shells before swallowing is necessary for proper utilization by C. carpio (Graveland et al., 1994). As gastropods have poor locomotive ability, development of higher strength shells will function as a defense strategy (e.g., Jordaens et al., 2006).
In the present study we explore the potential differences in predation ability of transgenic and nontransgenic C. carpio when fed different species and sizes of gastropods. Three predation experiments were conducted: 1) comparison of the maximum handling size when fed Bellamya aeruginosa of increasing sizes; 2) comparison of size selectivity when fed B. aeruginosa of different sizes; 3)comparison of species selectivity when co-fed B. aeruginosa and Radix auricularia, or Parafossarulus sinensis and Alocinma longicornis.
2 MATERIAL AND METHOD
2.1 Experimental fish and gastropods
The experimental fish were 1- and 2-year-old transgenic (age 1, mean total length 31.1 cm, mean biomass 384 g; age 2, mean total length 42.9 cm,mean biomass 980 g) and non-transgenic (age 1,mean total length 30.2 cm, mean biomass 351.4 g; age 2, mean total length 42.2 cm, mean biomass 922 g)C. carpio. They were obtained from the Institute of Hydrobiology, Chinese Academy of Science. For more details about the source of the experimental fish see Zhong et al. (2012). Both genotypes were reared in outdoor pools (200 m2, water depth 2 m, water temperature 10–28°C, dissolved oxygen>6.0 mg/L).The number of transgenic C. carpio available at the desired body size was highly restricted. Consequently only 15 fish from each age and genotype were transferred to the indoor re-circulating system and kept in four circular fiberglass tanks (diameter 150 cm, volume 1 000 L). All fish were initially fed to satiation with commercial pellets (Coland Ltd.,Wuhan, China). Then, gastropods described below were supplied to acclimatize the fish to gastropod feeding.
Four gastropod species were used in the present study, B. aeruginosa, R. auricularia, A. longicornis and P. sinensis. They are all common in ponds and lakes of China and have different shapes and shell strengths (Zhu et al., 2013a). Bellamya aeruginosa and P. sinensis were collected from Niushan Lake(Hubei, China), A. longicornis was collected from Honghu Lake (Hubei, China) and R. auricularia was collected from pools in Institute of Fisheries, Wuhan Academy of Agricultural Science & Technology(Hubei, China). All gastropods were held in four circular fiberglass tanks (diameter 150 cm, volume 1 000 L) with some Ceratophyllum demersum and Hydrilla verticillata so that the gastropods could feed and settle, and the tanks were connected to a recirculating system ( flow speed 0.2 L/s, water temperature 26–28°C).
2.2 Experimental procedures
Three predation trials, maximum handling size trial, size selection trial and species selection trial,were conducted under laboratory conditions. All trials were conducted in plastic tanks (98 cm×76 cm×68 cm,400 L). After 24 h aeration, tap water was added to the tanks at a depth of 60 cm. One third of the volume of water (about 150 L) was exchanged per day. The tanks were aerated (dissolved oxygen 8.2–10.9 mg/L,pH 7.6–8.0) and covered with mesh to prevent the fish from escaping. No sediment was added into the tanks.The water temperature was 28.2±0.9 (mean±standard deviation, n=15) °C and light conditions 12 h:12 h(L:D).
2.2.1 Maximum handling size trial
The maximum handling size trial was designed to compare the maximum handling size of transgenic and non-transgenic carp when foraging on B. aeruginosa.As transgenic fish were generally larger than the nontransgenic, attempts were made to match the size of both genotypes by selecting individuals with comparable sizes whenever possible. Five fish were chosen for each age and genotype (Table 1). The fish were transferred to the tanks (one fish per tank) and acclimated to the conditions over 3 days during which they were fed with 5 small (shell height 5–7 cm)B. aeruginosa. All fish were then starved for 24 h. The first trials started at 9:00 on the following day. For the 1-year-old group, each fish was presented with 17 B.aeruginosa (22.8 ind./m2) ranging from 5 to 22 mm in shell height (one snail for each 1-mm interval) and allowed to feed for 48 h. Uneaten snails were collected and measured to identify maximum handling size. For the 2-year-old group, each fish was presented with 13 B. aeruginosa (17.5 ind./m2) ranging from 11 to 23 mm in shell height. Different prey densities between 1-year-old group and 2-year-old group was due to limited number of available small size B. aeruginosa.Four small size B. aeruginosa that de finitely can be handled by 1 year group were removed in the trials of 2 year group to guarantee the supplement of small B.aeruginosa in the following trials. As there was no sediment added into the tanks, using different densities of prey could hardly in fluence the success of finding prey. All trials were repeated the following day.
2.2.2 Size selection trial
The size selection trial was designed to determine size preference of B. aeruginosa in transgenic andnon-transgenic C. carpio. As the transgenic fish are very expensive and rare, and the maximum handling size of 1-year-old fish was limited, only three 2-yearold fish for each genotype were used. All trials were carried out on single fish (one fish per tank) using the same experimental devices as in the maximum handling size trials. The fish were transferred to the tanks and acclimated to the conditions over three days during which they were fed half rations of gastropods.The fish were then starved for 24 h. The first trial started at 9:00 on the following day, and the replicate trial at 15:00. The trials were repeated the following day giving at total of three replicates for each fish. In each trial, the fish were presented with a total of 15 B. aeruginosa (20.1 ind./m2) from the three size classes (shell heights 5–6 mm, 8–9 mm and 11–12 mm, 5 snails for each size class). The trials ended after 15 min or when all snails were eaten. Uneaten snails were collected and measured.
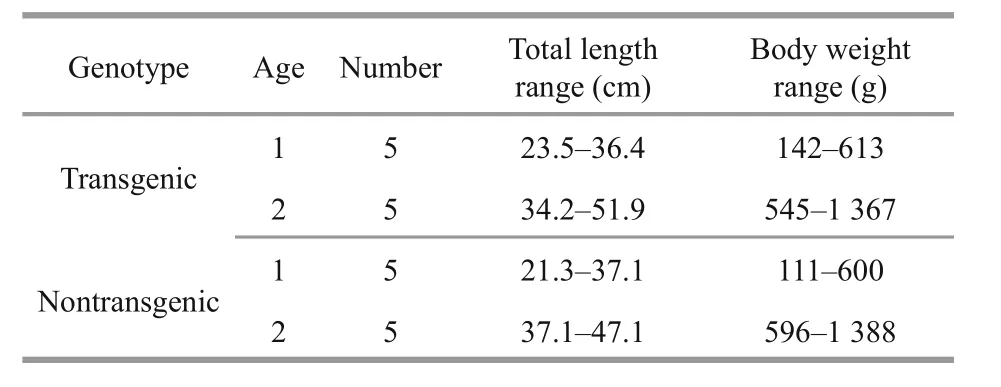
Table 1 Size of transgenic and non-transgenic C. carpio used in the maximum handling size trial
2.2.3 Species selection trial
The species selection trial was designed to determine species preference of transgenic and nontransgenic C. carpio on snail species. Only 2-year-old fish were used as the same reason described in the size selection trial. The snail species used here were B. aeruginosa, R. auricularia, A. longicornis and P. sinensis. The trials were divided into two subexperiments using the same experimental devices in the maximum handling size trials. The first subexperiment determined the species preferences between transgenic and non-transgenic C. carpio for B. aeruginosa or R. auricularia, as they differ markedly in shell strength. For these trials, three 2-year-old fish for each genotype were used. All trials were carried out on single fish (one fish per tank). The fish were transferred to the tanks and acclimated to the conditions over three days during which they were fed with gastropods at half satiation. All fish were then starved for 24 h. The first trial started at 9:00 on the following day, and the replicate trial at 15:00. The trials were repeated the following day giving at total of four replicates for each fish. Each fish was presented with 8 B. aeruginosa (10.7 ind./m2) ranging from 12 to 14 mm in shell height and 10 R. auricularia(13.4 ind./m2) ranging from 8 to 10 mm in shell height. Higher density in R. auricularia was to reduce the in fluence of prey size on the selection of experimental fish, as R. auricularia was limited resource and was generally smaller than B. aeruginosa.The trials ended after 15 min or when all snails were eaten. Uneaten snails were collected and measured.
The second sub-experiment aimed to determine species preference of transgenic and non-transgenic C. carpio fed two snail species with similar shell strength, A. longicornis and P. sinensis. The procedure was identical to the first sub-experiment except that five fish were used for each genotype and each fish was presented with 8 A. longicornis (10.7 ind./m2)ranging from 7 to 9 mm in shell height and 8 P.sinensis (10.7 ind./m2) ranging from 8 to 10 mm in shell height.
2.3 Data analysis
The maximum handling size was chosen as the largest prey eaten in the two replicate trials. Chesson’s selectivity index (Chesson, 1983) was used for all selectivity data, using the formula:
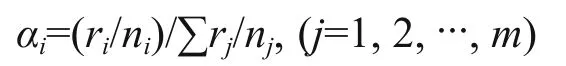
where, αiis selectivity index, the value ranges from 0(totally avoidance) to 1 (total selectivity), 0.5 means no selectivity (Chesson, 1983). For size selection trials, niis the number of B. aeruginosa in size class i given to each fish; riis the number of B. aeruginosa in size class i eaten by each fish; m is the number of size classes. For species selection trials, niis the number of snail species i provided to each fish; riis the number of snail species i eaten by each fish; m is the number of size species.
Linear regression of the maximum handling size on the size of experimental fish was used to investigate if the maximum handling size increased allometrically with fish size. Analysis of covariance (ANCOVA)was used to compare the difference in maximum handling size among genotypes from the same age group with fish total length as covariate. Mann-Whitney non-parametric test was used to compare the difference in maximum handling size among genotypes with both ages combined. Kruskal-Wallis test was used to compare the selectivity difference of each predator on prey size groups or species using repeated measurements data. The number ofgastropods consumed within a speci fic size class or species in all repeated measurements were pooled by predator’s genotype. Then a cross tabulated chisquare test was used to compare size and species selectivity differences among genotypes. Likewise,the individual differences of size and species selectivity within each genotype were compared. The statistical software was SPSS 13.0 (Systat Inc.,Chicago, IL, USA). Signi ficant differences were accepted at P <0.05.

Table 2 Linear functions between the maximum handling size (shell height, H S, mm) and the size (total length, L T, cm) of transgenic and non-transgenic C. carpio from 1- and 2-year-old groups when foraging on B. aeruginosa
3 RESULT
3.1 Maximum handling size
No signi ficant difference was found in the maximum handling size on B. aeruginosa among transgenic and non-transgenic carp. Using fish total length as covariance, the results of ANCOVA found no signi ficant differences among genotypes in both 1-year-old ( F= 0.734, d.f.=1.8, P=0.420) and 2-yearold groups ( F=2.108, d.f.=1.8, P=0.190). Mann-Whitney U test also showed no difference in the maximum handling size among genotypes ( U= 42.000,W= 97.000, P= 0.545) with both ages combined.
The maximum handling size on B. aeruginosa was positively correlated to fish size (Table 2). In the 1-year-old group, the relationships between maximum handling size and fish size resembled a J-shaped curve where the largest transgenics (total length LT=36.4 cm)and non-transgenics ( LT=37.1 cm) could eat B. aeruginosa at sizes 15.9 mm and 20.6 mm,respectively (Fig.1). In the 2-year-old group, the relationships between maximum handling size and fish length resembled an S-shaped curve with the in flection point to fish size being about 41.3–47.7 cm in transgenics and 43.7–47.1 cm in non-transgenics.The largest transgenics ( LT=51.9 cm) and nontransgenics ( LT=47.1 cm) could eat B. aeruginosa at sizes 18.3 mm and 22.0 mm, respectively.
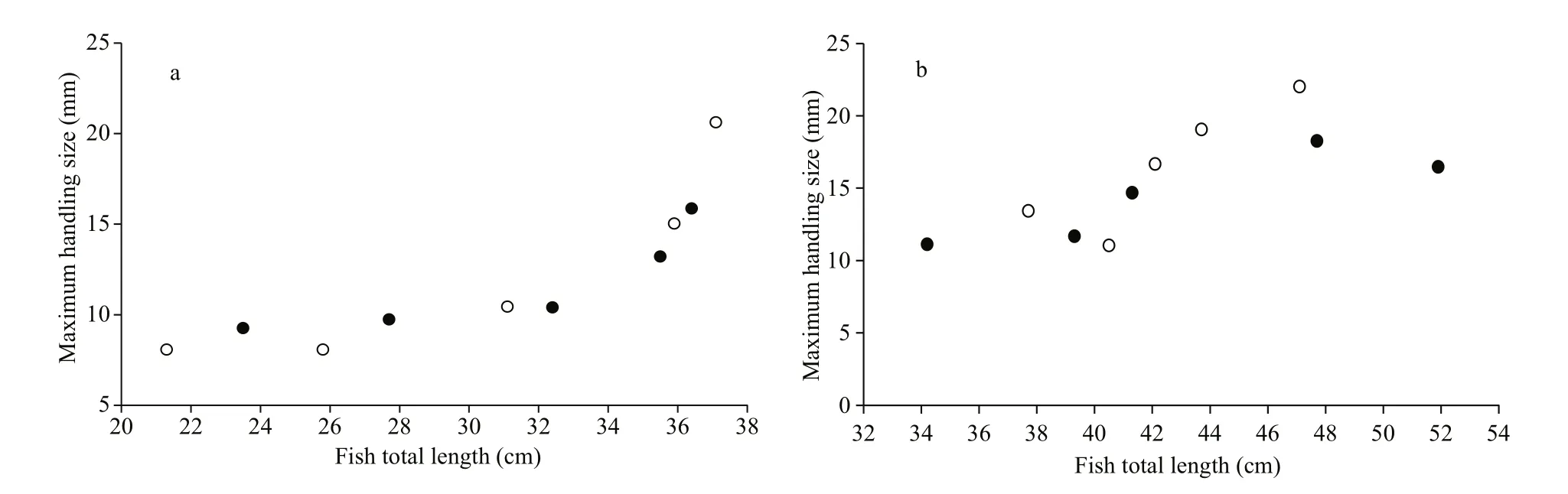
Fig.1 The maximum handling size of transgenic (black) and non-transgenic C. carpio (white) when foraging on B. aeruginosa
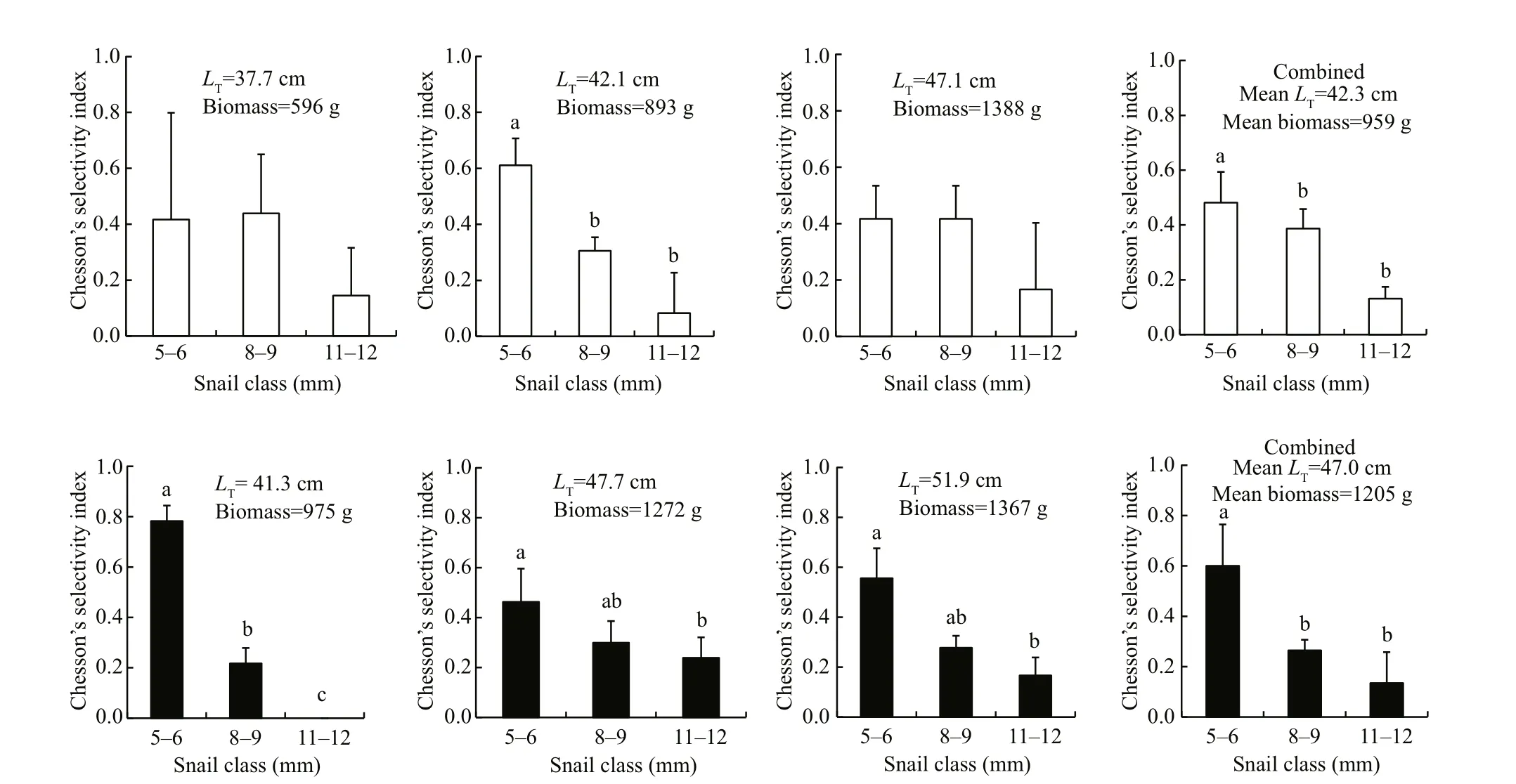
Fig.2 Chesson’s selectivity index (±SD) of 2-year-old transgenic (black) and non-transgenic C. carpio (white) when foraging on B. aeruginosa with different sizes
3.2 Size selection
There were no signi ficant difference in prey size preference between two genotypes ( χ2=0.941,P= 0.625). There was also no signi ficant individual difference within the transgenics ( χ2=7.608, P=0.107)and within the nontransgenics ( χ2=2.775, P= 0.596).Both transgenics and non-transgenics showed signi ficant size selectivity for B. aeruginosa (Fig.2).The individuals of the small-sized class were selected while the individuals of the large-sized class were avoided for both genotypes.
3.3 Species selection
When given the option to choose between B. aeruginosa and R. auricularia, both transgenics and non-transgenics favored R. auricularia with no signi ficant difference in preference between two genotypes ( χ2=2.257, P=0.133) (Fig.3). There was also no signi ficant individual difference within the nontransgenics ( χ2=5.100, P= 0.078). While signi ficant individual difference was found within the transgenics ( χ2=7.028, P= 0.030).
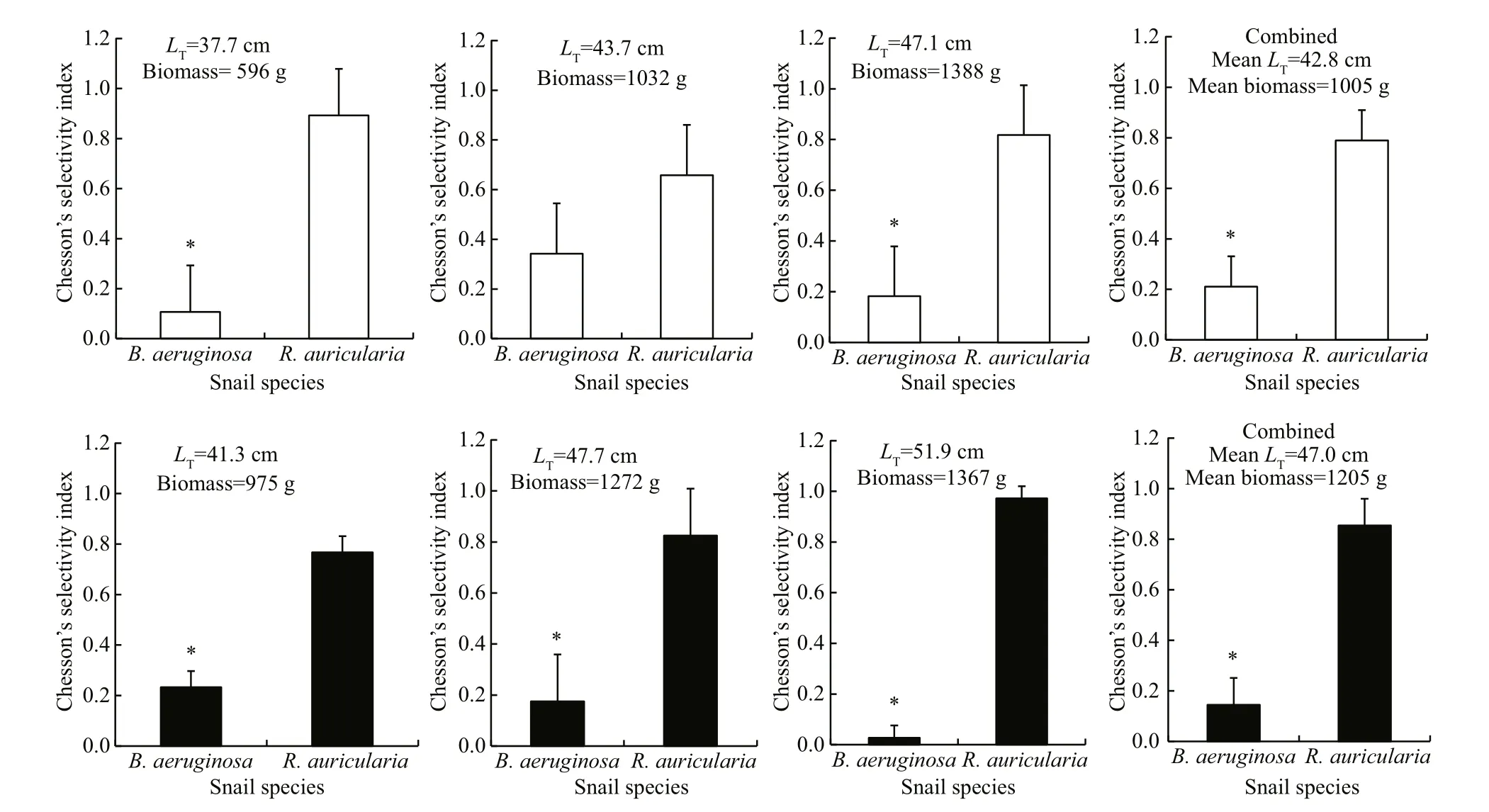
Fig.3 Chesson’s selectivity index (±SD) of 2-year-old transgenic (black) and non-transgenic C. carpio (white) when foraging on B. aeruginosa and R. auricularia with similar individual size
Likewise, when given the option to select between A. longicornis and P. sinensis, both genotypes displayed no signi ficant preferences ( χ2=1.125,d.f.=1.8, P=0.289) (Fig.4). There was also no signi ficant individual difference within the transgenics( χ2=3.990, P= 0.407) and within the nontransgenics( χ2=1.916, P= 0.751).
4 DISCUSSION
Due to a low number of available experimental fish, only few fish were used in the present study, and the results were preliminary. However, our study provided the first report of selective predation performance of transgenic carp, and may have a certain reference value for future studies.
Maximum handling size re flects the feeding ability of fish. Larger individuals thus have better feeding ability (e.g., García-Berthou, 2001). In this study,there was also a positive relationship between total length of fish and their maximum handling size for both transgenic and non-transgenic C. carpio.Differences in mouth/pharyngeal physiology may in fluence molluscan feeding ability between different molluscivorous fish. Meyer (1989) found the feeding ability of two cichlids Amphilophus citrinellus on two snails with different shell strength correlated to the morphology of pharyngeal teeth. The crushing ability of redear sun fish, Lepomis microlophus feeding on snails was more than 2 times higher than that of pumpkinseed sun fish, Lepomis gibbosus, while the feeding ability of small L. microlophus on snails was limited by the bite force (Huckins, 1997). In the present study, there was no signi ficant difference in maximum handling sizes between two genotypes,even though transgenic fish showed relatively small maximum handling size compared to similar nontransgenic fish. This deviated more or less from our prediction that the maximum handling size consumed by transgenic fish is signi ficantly lower than the nontransgenic, because of the delayed or impaired pharyngeal bone development in transgenic fish (Zhu et al., 2013b). There would be two possible reasons for this discrepancy. Firstly, the difference in pharyngeal bone development in our experimental fish with considerable size may be too small to cause difference in feeding ability. Secondly, transgenic C. carpio may compensate for delayed pharyngeal bone development in other ways. For example,pharyngeal jaw force in C. carpio is insufficient to crush some hard preys, but they can therefore transfer large forces from the dorsal and ventral trunk muscles to the pharyngeal jaw by lifting the skull and retracting the pectoral girdle to enhance the pharyngeal jaw force (Sibbing, 1988). Transgenic C. carpio has much thicker dorsal trunk muscle (Wang et al., 2001), which would compensate for reduced pharyngeal strength.
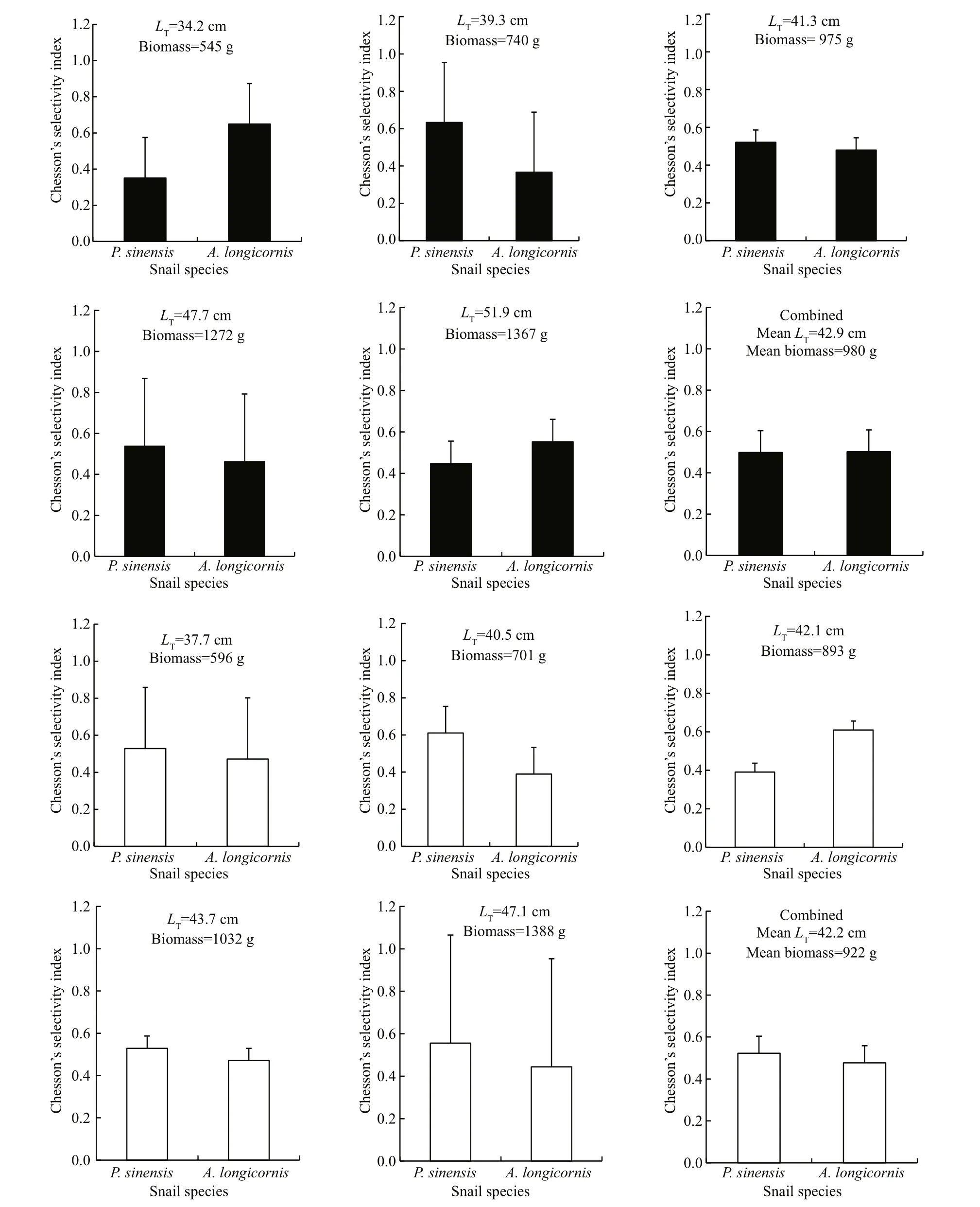
Fig.4 Chesson’s selectivity index (±SD) of 2-year-old transgenic (black) and non-transgenic C. carpio (white) when foraging on P. sinensis and A. longicornis with similar individual size
Many factors like genetic background, body size,hunger, size, species and density of prey would in fluence the prey selectivity. Size plays an important role in the prey-predator relationship (Lundvall et al.,1999). Large prey can be found easier (Pennings,1990), but large prey are harder to handle, and need more time and energy to consume. Therefore,predators need to balance the bene fit to cost before choosing prey size and species. Previous data have shown that C. carpio prefer V. piscinalis with small,thin and easier crushed shells than P. annulata with thick shells (Covich and Knezevic, 1978). Our study also showed similar results in both transgenic and non-transgenic fish. In the size selection trials, small B. aeruginosa was the favorite prey, while large B. aeruginosa with thick shell was avoided. In the first species selection experiment, R. auricularia with thin shells was favored by the predators, while B. aeruginosa with thick shell was avoided. In the second species selection experiment, however, the predators had no preference when fed two gastropod species with similar shell strength. Therefore, the result indicates that both transgenic and nontransgenic carp prefer smaller and easier crushed gastropods.
Previous studies have found that size distribution and reproduction patterns (Johnson and Belk, 1999),community structure (Zaret, 1980; Brönmark, 1988),food web stability and species composition (Faria and Da Silveira Costa, 2009) can be in fluenced by prey selection. Negative correlation was found between the density of C. carpio and the diversity of macroinvertebrate (Parkos III et al., 2003), and C. carpio can affect the diversity of macrozoobenthos community directly by predation (Zambrano and Hinojosa, 1999). In the present study, both transgenic and non-transgenic C. carpio had similar upper prey size limits when fed B. aeruginosa. Furthermore,there were no signi ficant differences in size and species selectivity between transgenic and nontransgenic C. carpio with at least 1 year old fish (total length more than 37 cm). These results suggest that the effect of release or escape of transgenic C. carpio on the size and species structure of gastropods may be similar to that of wild C. carpio given no behavioral difference in locating the gastropods among transgenic and non-transgenic C. carpio. However, it would be necessary to conduct veri fication experiments under semi-natural or natural conditions. In addition, the present study may be of little value to estimate the in fluence of transgenic C. carpio on other macroinvertebrate species without hard shells, as the predation ability of transgenic C. carpio would not be restricted by its impaired pharyngeal bone development. Therefore, the future studies should also focus on the behavioral difference between the transgenic and non-transgenic C. carpio in locating food items and food intake under natural conditions,which may be of great signi ficance for the risk assessment of transgenic C. carpio on its prey.
5 CONCLUSION
Our study revealed that maximum size of B. aeruginosa consumed by transgenic C. carpio was slightly smaller than non-transgenic congener, but there was no signi ficant difference in maximum handling size between the two genotypes. The maximum handling size was positively correlated to the individual size of C. carpio. Both transgenic and non-transgenic C. carpio preferred small or thinshelled gastropods. No signi ficant differences in size and species selectivity on gastropods were found between the two genotypes. These results suggest that the potential effect of transgenic C. carpio on the size and species structure of gastropods may be similar to that of wild C. carpio.
6 DATA AVAILABILITY STATEMENT
All data supporting the findings of this study are available within the article.
猜你喜欢
杂志排行

Journal of Oceanology and Limnology的其它文章
- Editorial Statement
- Recent insights into physiological responses to nutrients by the cylindrospermopsin producing cyanobacterium,Cylindrospermopsis raciborskii*
- Response of Microcystis aeruginosa FACHB-905 to different nutrient ratios and changes in phosphorus chemistry*
- In fluence of light availability on the speci fic density, size and sinking loss of Anabaena flos- aquae and Scenedesmus obliquus*
- Application of first order rate kinetics to explain changes in bloom toxicity—the importance of understanding cell toxin quotas*
- Regime shift in Lake Dianchi (China) during the last 50 years*