现代舞训练与弦乐训练对脑灰质体积的差异影响*
2018-07-07李谷静尧德中
李谷静 李 薪 贺 辉 罗 程 尧德中
(电子科技大学生命科学与技术学院, 成都 610054)
1 引言
大脑的可塑性受多种因素影响, 其中长期学习训练是其结构改变的重要因素(Draganski et al.,2004)。舞蹈或音乐训练是指依据科学原理, 计划性地对受训者进行机体结构机能重建和技术加工,使其通过长期练习掌握常人不易获得的特殊技能的过程(覃嫔, 2018; 马清, 2000)。两类训练的特殊性与长期性为研究脑可塑性提供了良好模型。舞蹈训练与音乐训练具有许多异同点:两者都是复杂感觉运动过程, 涉及大脑对多种感知觉信息的加工整合; 都是情感表达性艺术训练, 需要脑对内部躯体变化与外部刺激信息进行交互加工, 实现艺术情感的表现。不同在于舞蹈训练是以舞者自身为工具的韵律性动觉训练, 它强调本体感觉与身体控制, 强调视−听−动的高度整合。舞蹈学习往往通过大量的动作观察与动作模仿而实现(平心, 2004)。音乐训练特别是器乐训练, 是以掌握特殊演奏技能为目标的训练, 它强调听觉与手指精细运动的高度协调与快速反馈, 要求听−动的高度整合。音乐学习通过听觉信息修正动作,从而达到对演奏技能的修正(Mutschler et al., 2007;蒋存梅, 2016)。
目前, 相关研究主要从脑功能与脑结构的改变来刻画两种训练的可塑性影响。关于舞蹈的研究主要聚焦于动作观察网络(action observation network, AON)以及感觉运动网络。研究AON主要是为探索舞蹈专业性对动作知觉以及对目标导向性行为的影响。这些研究发现个体通过舞蹈训练所获得的真实动作经验, 与动作观察所获得的间接经验都会影响其动作观察网络, 具体来讲两种动作经验都会增进该网络的激活(Calvo-Merino,Glaser, Grèzes, Passingham, & Haggard, 2005; Cross,Hamilton, & Grafton, 2006; Jola et al., 2013)。与此同时, 研究者们通过扫描舞者执行下肢舞蹈动作时的脑功能活动情况, 进而研究舞蹈动作中独特的感觉运动过程。其结果显示舞蹈动作执行时与本体感觉、动作控制、节奏保持等功能相关的皮层、皮层下结构及小脑等广泛区域被激活(Brown,Martinez, & Parsons, 2006; Ono et al., 2014)。关于脑结构方面的研究也表明长期舞蹈训练将导致感觉运动区域的灰质与白质结构发生广泛改变。Hänggi, Koeneke, Bezzola 和 Jäncke (2010)首次开展芭蕾舞蹈家的大脑结构研究。他们发现舞蹈家的左侧前运动皮层、辅助运动皮层、壳核与额上回的灰质体积显著降低; 双侧皮质脊髓束、双侧内囊与胼胝体的白质体积显著降低; 连接前运动皮层的白质纤维微结构出现各向异性降低。舞蹈家大脑的广泛结构改变引起了其他研究者的关注。Giacosa, Karpati, Foster, Penhune和 Hyde(2016)通过进一步对比舞蹈家与音乐家的白质纤维微结构, 发现舞蹈家感觉运动网络内的白质纤维结构(如皮质脊髓束、上纵束和胼胝体)变化明显。较之音乐家与对照组, 舞蹈家的白质纤结构更为发散和长程, 连接了更为广泛的感觉运动皮层。综上所述, 舞蹈训练对脑结构的特异性影响主要表现在涉及感觉运动的广泛脑区发生改变,具体表现为功能皮层灰质体积的减小, 且连接这些皮层的白质纤维微结构发生改变(各项异性降低)。关于音乐的研究主要集中于听−动整合相关网络以及音乐认知的神经过程。研究发现音乐家的感觉运动皮层、听觉皮层及小脑的功能增强,说明音乐训练可能提升了大脑对听−动信息的处理与整合(Baumann et al., 2007; Bangert et al.,2006)。另一类研究则揭示音乐训练中的句法加工、工作记忆和音乐情绪加工的神经过程, 结果发现音乐训练增强了额下回、顶下小叶、脑岛等相关脑区的功能(Groussard et al., 2010; Koelsch &Siebel, 2005)。关于音乐家脑结构的研究结果基本与其脑功能的特异性改变一致。相关研究发现灰质体积的增加与音乐专业性呈显著正相关(Bermudez,Lerch, Evans, & Zatorre, 2009), 且分布在与音乐技能训练高度相关的脑区, 如颞横回(Schneider et al., 2002)、左侧颞平面(Schlaug, Jancke, Huang, &Steinmetz, 1995)、颞上回(Bermudez et al., 2009)、中央沟内手指感觉区域(Li et al., 2010)、小脑(Hutchinson,Lee, Gaab, & Schlaug, 2003)等。也有研究发现音乐家的手指运动区(位于中央前回)的灰质结构更清晰和典型(Bangert & Schlaug, 2006)。相应的, 音乐家大脑中连接以上灰质区域的白质结构也发生特异性改变, 出现各项异性的增高, 如连接前运动皮层与感觉运动皮层的胼胝体前部和后部(Öztürk,Tasçioglu, Aktekin, Kurtoglu, & Erden, 2002)、连接皮层−脊髓的皮质脊髓束(Rüber, Lindenberg, &Schlaug, 2015)等。总的来说, 音乐训练对脑结构的特异性影响主要表现在与听觉、手指运动与语义分析高度相关的皮层发生改变, 具体表现为皮层灰质体积的增大, 且连接这些区域的白质纤维微结构发生改变(各项异性增高)。
综上所述, 舞蹈与音乐的脑可塑性研究呈现以下几点不足:对音乐的关注远远多于舞蹈, 舞蹈的相关研究还有待于进一步丰富与深入。结构研究远远少于功能研究, 且有限的结构研究结果并不稳定。单独研究舞蹈或音乐的可塑性改变较普遍, 而比较两种训练导致的差异非常有限, 仅有两篇文章报道了舞蹈家与音乐家的白质与灰质结构的异同(Giacosa et al., 2016; Karpati, Giacosa,Foster, Penhune, & Hyde, 2017)。对两种训练导致的灰质体积变化结果存在争议, 如 Giacosa等人(2016)发现舞蹈家与音乐家灰质体积在颞叶上部有共同的增加, 但最近发表的一项研究则报告舞蹈训练对灰质结构的改变并无显著影响(Burzynska,Finc, Taylor, Knecht, & Kramer, 2017)。除了上述不足, 我们的前期研究结果还无法完整描述舞蹈训练的特异性影响。我们虽然发现舞蹈训练对大脑皮层−基底节回路的连接具有增强作用(Li et al.,2015), 说明舞蹈训练提高了受训者的感觉运动整合功能, 但这种改变也发生在全身性运动的体育训练中(Huang et al., 2017)。同时由于舞蹈与音乐关系最为密切, 它往往在音乐节奏中进行, 因此要刻画舞蹈的特征就涉及对音乐效应的考虑。所以我们将研究重点放在两种训练的脑结构对比上,希望一定程度上分离舞蹈中的音乐影响, 进一步探索舞蹈训练相关的脑结构特异性改变。基于前述的研究概况与深入探索舞蹈特征的需要, 我们选取接受现代舞与弦乐训练的被试, 对两种被试的脑灰质体积进行比较研究。选择现代舞是因为它更接近舞蹈本质。相对于单纯追求形式与技巧的芭蕾, 现代舞更强调人体动作本身。这正如表现主义现代舞创始人玛丽.魏格曼在关于舞蹈本体论中所述, 舞蹈的核心因素是人体动作, 即人体动作构成的“力”的因素, 因此没有音乐要素的舞蹈也能独立存在(吕艺生, 2013)。选择弦乐是因为它更典型地体现了音乐训练中的听−动整合。相对于吹奏类与键盘类乐器的训练, 弦乐训练对演奏者音准要求非常高。弦乐演奏音高的变化是演奏者根据听觉反馈, 精准确定手指在琴弦上的位置而实现(马清, 2000)。因此弦乐训练涉及快速、密集和精确的听−动整合。
本文的创新与意义在于:训练组的类型齐整统一。我们选取相同舞蹈风格与乐器类型的被试,在脑结构可塑性分析研究中降低了舞蹈训练风格与乐器不一致因素的干扰, 进而更好地描绘两种训练对脑结构的特异性影响。研究的问题具有探索性。关于两种训练对灰质结构影响的研究非常有限且存在争议, 我们的研究能为这一问题提供新的证据。研究具有延续性。基于课题组前期研究发现, 我们通过比较进一步刻画舞蹈训练对脑结构的独特影响。
本研究采用基于体素的形态学分析方法(voxelbased morphometry, VBM)进行组间灰质体积对比。我们预测训练组的脑灰质体积可能存在如下改变:现代舞与弦乐训练组的变化范围主要位于感觉运动网络; 现代舞训练组可能出现灰质体积的减小, 而弦乐训练组则可能是增加; 两组被试还可能出现颞叶上部灰质体积的共同增高。
2 材料与方法
2.1 研究对象及纳入标准
本研究招募了3组被试, 包括18名现代舞被试(现代舞训练组)、20名弦乐被试(弦乐训练组)及25名对照被试(对照组) (见表1)。训练组被试均为大学艺术特长生或艺术专业学生, 其中现代舞训练组主修现代舞, 弦乐训练组主修西洋弦乐(小提琴、中提琴或大提琴)。在纳入本实验前, 两组专业被试分别接受两名舞蹈或两名音乐专业教师的综合评估, 其专业性被评价为达到舞蹈专长或音乐专长条件。舞蹈教师对现代舞训练组被试进行如下评估:舞蹈模仿。向被试示范3次长度为 4个八拍的全新舞蹈动作(被试之前从未训练过), 要求被试根据自己的记忆重复舞蹈片段, 评估其动作完整性与准确性。舞蹈基本功。要求被试完成下横竖叉、下腰等动作, 同时要求被试根据自己的软开度进行跳、转、翻的技巧组合, 评估其柔韧度与技巧。舞蹈即兴。要求被试根据随机选择的音乐进行即兴舞蹈, 评估其舞蹈动作运用与舞蹈表现力。音乐教师对弦乐训练组被试进行如下评估:自选曲目演奏。要求被试演奏一首自选曲目(3分钟以内), 考察被试的器乐演奏与表现力。视奏能力。音乐教师谱写一段16个小节的中等难度的演奏片段(被试之前从未演奏或练习过), 要求被试现场演奏, 考察其识谱演奏能力。现代舞训练组被试均无音乐训练经历, 弦乐训练组被试均无舞蹈训练经历。两组专业被试训练年限从7至17年不等。对照组为与之匹配的大学生,无任何舞蹈、音乐训练经验。现代舞训练组、弦乐训练组及对照组在年龄、性别和受教育年限方面均无统计显著性差异, 两种训练组在训练年限方面无统计显著性差异, 分组达到均衡性(见表1)。3组被试完成艾丁伯格手性问卷(Oldfield, 1971)测试, 均为右利手。所有被试均无神经系统疾病、发展性疾病或物质滥用。本实验符合赫尔辛基宣言, 并获得作者所在研究机构伦理委员会许可,所有被试签署实验知情同意书。
2.2 数据获取
本研究在电子科技大学信息医学中心完成。MRI数据的采集设备为GE Discovery 750 3.0T磁共振系统(GE Medical Systems, Milwaukee, WI),使用标准的GE全头线圈。高分辨率T1加权图像通过3D梯度回波序列(T1-3DFSPGR)获得。扫描参数为:TR = 6.008 ms, TE = 1.984 ms, 反转角 9˚,矩阵 256×256, FOV=25.6 cm × 20 cm, 层厚 1 mm,层数 152层。采集数据时, 被试在扫描仪中尽量保持头部不动。实验使用海绵来固定被试头部以减少头动, 使用耳塞来降低扫描过程中的噪音。
2.3 数据处理及统计分析
VBM是一种全脑无偏算法, 用来计算脑体积与容量(Ashburner & Friston, 2000, 2005)。本研究采用VBM8工具包进行灰质分析。(1)首先对3组被试的T1加权脑图像进行数据预处理, 包括检查

表1 被试人口学信息
所有被试数据的成像质量, 手动调整所有结构 MRI的原点到前连合。(2)其后在 MATLAB 2014环境下使用嵌套于SPM8 (http://www.fil.ion.ac.uk/spm)内的VBM8工具包对被试三维T1加权脑图像进行常规的灰质体积估计流程:1)将所有被试的结构MRI图像分割成灰质、白质和脑脊液。2)使用DARTEL算法对分割后的灰质图像建立基于组水平的模板。3)利用构建的模板将所有灰质图像配准到蒙特利尔神经研究所(Montreal Neurological Institute, MNI)标准人脑模板空间。4)将配准后的图像进行调制得到灰质体积图像。5)使用半高全宽为 8毫米高斯核将配准后的灰质图像进行平滑。(3)最后对3组被试平滑后的灰质MRI图像进行组间单因素方差分析(进行高斯随机场理论校正, 校正后p< 0.05) (Nichols, 2012), 进而对差异脑区通过两两比较进行事后检验分析(双样本t检验,p< 0.05)。
3 结果
较之对照组, 现代舞训练组与弦乐训练组均表现出灰质结构的特异性改变(见表2)。其中现代舞训练组的双侧丘脑、右侧辅助运动皮层(SMA)与左侧中央前回(M1)灰质体积显著低于对照组和弦乐训练组; 其右侧眶部额上回、右侧壳核与左侧小脑则显著高于其他两组(p< 0.05,t检验) (见图1)。弦乐训练组的右侧额中回、右侧颞上回与左侧中央前回灰质体积显著大于对照组和现代舞训练组(p< 0.05,t检验) (见图 1)。

表2 现代舞训练组、弦乐训练组与对照组灰质体积的组间比较
4 讨论
VBM分析结果表明, 较之对照组现代舞训练组与弦乐训练组的灰质体积存在特异性改变, 两者灰质体积改变的区域和方式存在差异。现代舞训练组的改变涉及左侧小脑、双侧丘脑、右侧壳核、右侧辅助运动区等有关感觉运动控制的广泛脑区, 不同区域灰质体积既有增加也有减小。弦乐训练组的改变集中于听−动−读相关脑区, 并统一表现为增加的灰质体积。组间灰质体积的特异性改变说明现代舞训练与弦乐训练对脑可塑性的不同影响。
4.1 现代舞训练对脑灰质体积的影响
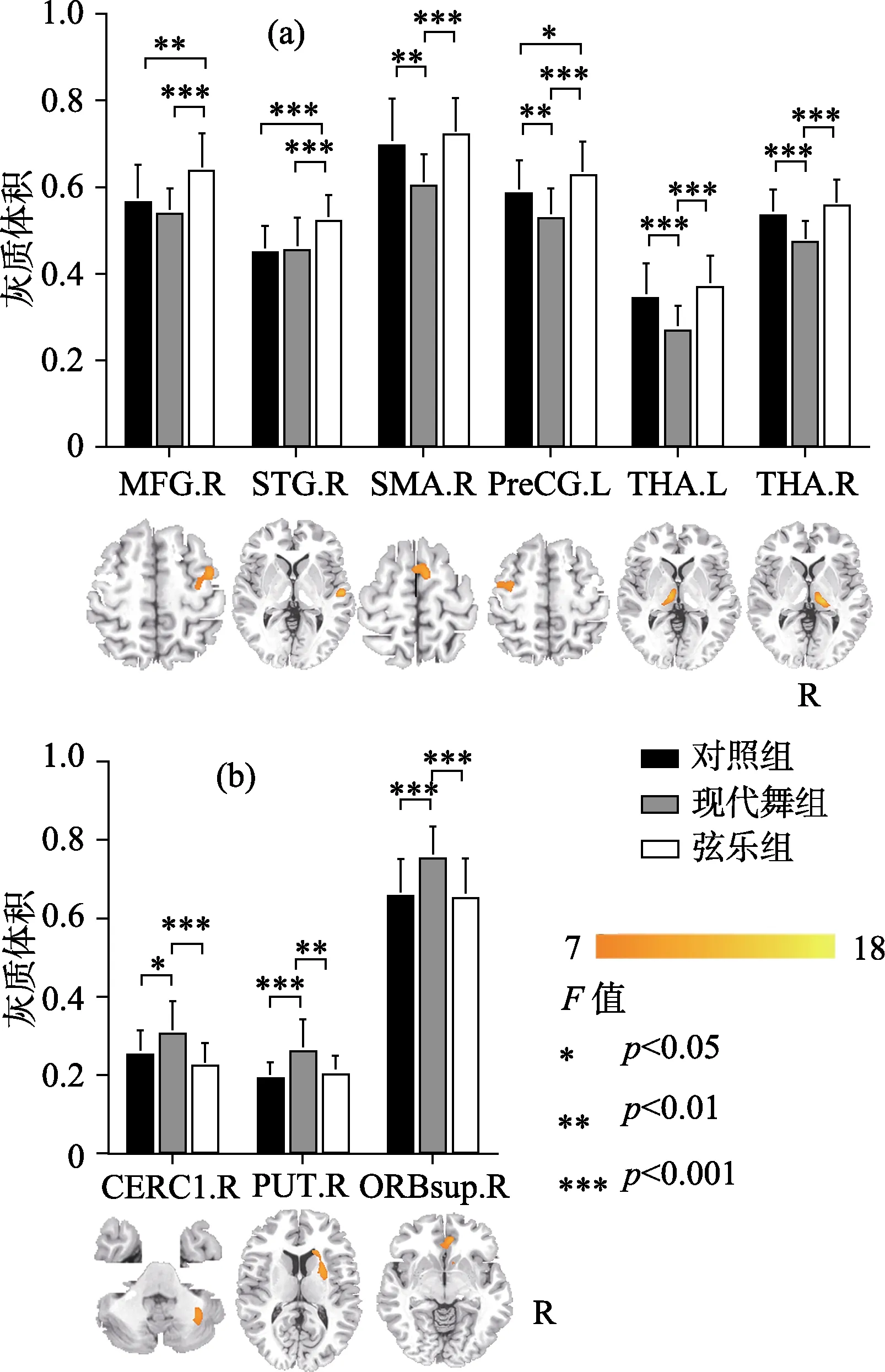
图1 现代舞训练组、弦乐训练组与对照组灰质体积的组间比较
较之弦乐训练组与对照组, 现代舞训练组大脑灰质体积变化区域更广泛, 在这些区域中同时存在灰质体积的增高与降低。该结果可能反映了全身性舞蹈动作训练对脑的特异性影响。Brown等人(2006)利用正电子发射断层成像(positron emission tomography, PET)技术观察到舞者进行下肢舞步移动时的脑功能状态, 结果发现小脑蚓部、右侧壳核、腹侧丘脑、内侧上小叶、辅助运动皮层(SMA)、初级运动皮层(M1)与前运动皮层区域的激活明显。我们的结果基本属于以上功能区内,从结构上说明舞蹈训练可能影响感觉运动网络内的广泛脑区。另外, 现代舞训练组灰质体积的增减也可能体现某种结构与功能的优化。增高的灰质体积往往意味着该区域的神经元更多, 处理信息的效率更高, 对应的认知和行为功能更强大。大部分相关研究表明训练水平往往与灰质体积成正比(Maguire et al., 2000)。与此一致, 现代舞训练组在动作控制相关脑区中表现出增高的灰质体积:比如眶部额上回, 该区域与身体平衡维持相关(Taubert et al., 2010); 小脑下部, 该区域将本体感觉信息与运动前庭功能相整合, 与平衡保持及姿势维持高度相关(Kheradmand & Zee, 2011)。现代舞训练要求舞者不断创造自发动作, 很多自发动作的组合可能是全新的, 因此需要大脑付出更多努力来进行各种动作和姿态的调控与保持, 进而导致以上两个区域灰质体积的增大。其次现代舞训练组还表现出右侧壳核灰质体积的增高与双侧丘脑灰质体积的降低。这一相反变化可能与壳核与丘脑在节奏运动中的功能有关。相关研究报道舞蹈家的右侧壳核在节奏运动条件下激活明显,而丘脑在非节奏复杂运动时有明显激活(Brown et al., 2006)。我们前期的研究也发现舞蹈家的壳核与全脑的功能连接更显著(Li et al., 2015)。现代舞训练强调动作性, 常常通过动作的力度变化来表现舞蹈情感, 而这种力度变化往往借助于动作节奏的改变而实现。因此, 在现代舞训练中不仅需要舞者保持动作的节奏性, 还需要在不同节奏间进行动作切换。这必然需要壳核的高度参与而丘脑保持抑制, 因此可能导致这两个区域相反的灰质体积变化。进一步我们还发现现代舞训练组右侧辅助运动皮层(SMA)与左侧初级运动皮层(M1)的体积显著减小。这两个脑区在复杂运动控制和感觉运动表征方面扮演重要角色, 是参与舞蹈过程的重要脑区。关于芭蕾舞蹈家和体操运动员的相关研究均发现这两个脑区的灰质体积显著降低(Hänggi et al., 2010; Huang, Lu, Song, & Wang,2015), 这可能体现了全身性运动技能训练的特殊效应。综上所述, 现代舞训练所影响的灰质结构变化范围更广泛, 灰质体积增高与降低的区域并存。同时这些变化都分布于皮层−基底节−丘脑−小脑神经回路内(该回路贯穿全脑, 与高级认知、运动调控高度相关) (Bostan, Dum, & Strick, 2013)。以上结果可能反映了现代舞独有的训练效应。相对于注重形式和规范的传统舞蹈, 现代舞是一种更为自由和灵活的舞种, 强调创造性的自发动作,要求舞者容纳和表现更为复杂和广泛的内外感受。这一特点可能需要脑在更大尺度上去加工和整合信息。因此我们推测现代舞训练影响的可能是涉及全脑感觉运动功能协同的广泛脑区。
4.2 弦乐训练对脑灰质体积的影响
较之对照组和现代舞训练组, 弦乐训练组灰质体积的改变集中于与音乐技能直接相关的脑区,且全部是增高。这一发现显示弦乐训练这种听−动结合的精细运动对大脑的特异性影响。听是音乐家的重要能力之一。我们的结果表明弦乐训练组在右侧听皮层(颞上回)有显著的体积增高。相关元分析研究显示, 颞上回在音乐训练中协调听觉系统对错误的检查, 负责对音乐旋律的解码和音乐语义的记忆并参与音乐中的动−听整合过程(周临舒, 赵怀阳, 蒋存梅, 2017; Lahav, Saltzman, &Schlaug, 2007)。此外大脑听皮层对音乐的处理具有某种程度的偏侧性, 其中右侧颞上回对音乐更为敏感(Hyde, Peretz, & Zatorre, 2008)。相关的结构研究也发现音乐家颞上回的灰质体积显著增大,其体积与音乐训练时间成正比(Groussard et al.,2014)。除了听觉皮层的变化, 我们发现弦乐训练组左侧初级运动皮层(M1)的灰质体积显著增加。这个区域是所有运动执行的核心区域, 类似的结果也出现在各类器乐演奏家的灰质结构研究中(Bermudez et al., 2009; Han et al., 2009; Sluming et al., 2002)。进一步我们还发现弦乐训练组右侧额中回灰质体积的增加。这个区域与工作记忆的处理与存储相关(Jones, Adlam, Benattayallah, & Milton, 2017), 且与颞中回共同完成语义启动(Laufer, Negishi, Lacadie,Papademetris, & Constable, 2011)。相关研究发现音乐家在这一部位的灰质体积是增加的(Bermudez et al., 2009)。弦乐训练需要受训者识别乐谱符号并记忆乐谱, 这个过程会频繁调用语义表征与工作记忆的脑区, 可能促成该脑区灰质结构的特异性改变。综上所述, 弦乐训练对皮层的影响可能更为集中, 其灰质体积变化区域位于与训练高度相关的听觉、运动与语义加工皮层。这些区域表现出灰质体积的一致增高。我们推测这种改变可能与弦乐训练要求的高度精准性相关。相对于其他乐器训练, 弦乐训练需要更为细微精准的手指控制和稳定的内部音高知觉。受训者需要通过频繁的听−动信息整合与反馈, 才能进行正确演奏。因此弦乐训练可能导致与训练技能直接相关的皮层发生改变。
4.3 本研究的有趣发现
4.3.1 初级运动皮层
初级运动皮层(M1)负责发出运动控制指令及对运动进行表征, 它参与所有动作的执行, 是舞蹈与音乐训练相关的重要脑区。有趣的是, 我们发现现代舞训练组和弦乐训练组灰质体积在左侧M1出现了相反的改变趋势。现代舞训练组较之对照组与弦乐训练组灰质体积显著降低, 而弦乐训练组则显著增高。现代舞训练组 M1灰质体积的减小可能是神经修剪和优化的结果。通过科学专业的动作训练, 能让相关脑区内的神经元去除冗余突触, 增强局部回路内部以及远距离脑区间的连接, 从而建立更为高效的神经连接模式(段旭君,2013)。这一神经可塑性改变外化于行为就是自动化舞蹈动作的获得, 每个舞蹈动作调用较少的神经资源就可以完成。弦乐训练组M1灰质体积的增高则可能是弦乐高度特异性的技能所致。M1在手指或手部的有序运动时会被明显激活(Shibasaki et al., 1993)。同时该区域对运动频率非常敏感, 它的激活程度随着频率的增加而增大(Turner, Grafton,Votaw, Delong, & Hoffman, 1998)。弦乐训练是精细有序的手指运动, 要求给予高频快速的动作反馈, 这种训练方式可能会强烈地激活M1, 进而导致该区域灰质体积的增大。
4.3.2 颞上回
Karpati等人(2016)发现舞蹈家与音乐家灰质体积在这个区域有共同的增加, 并推测该结果与音乐舞蹈训练所涉及的大量动−听整合相关。我们的结果与他们相左, 仅在弦乐训练组中发现灰质体积的显著增高, 而现代舞训练组没有显著变化。其原因可能来自被试的差异。现代舞的典型特点是关注自发动作本身, 它并不高度依赖音乐线索, 甚至可以在无音乐的背景下进行。因此现代舞训练组涉及的动−听整合训练并不突出, 颞上回的灰质体积改变不显著。而弦乐训练则需要高度依赖听觉信息来精确调控手指动作, 更有力地训练了演奏者的动−听整合功能, 因此导致颞上回灰质体积的显著增高。
5 结论
本研究采用基于体素的形态学分析方法, 比较现代舞训练与弦乐训练对脑灰质体积的可塑性影响。结果表明, 两种训练都使脑灰质体积发生特异性改变。其中现代舞训练的影响区域更为广泛, 分布于皮层−基底节−丘脑−小脑回路内, 其变化既有增加也有减小。这一结果说明现代舞训练影响的可能是涉及复杂感觉运动整合的广泛脑区。弦乐训练影响的则是与训练特性(动−听)直接相关的脑区。通过比较研究, 我们的结果首次展示了现代舞训练与弦乐训练对脑灰质结构影响的差异, 揭示了舞蹈训练对脑的特异性影响, 进而为舞蹈治疗和音乐治疗提供神经学基础。此外,我们的结果也提示后续有两个问题需要进一步探索:其一, 长期舞蹈训练对关键脑区的影响方式是怎样的?我们发现现代舞训练组存在灰质体积增高的区域, 如小脑与壳核, 而大部分相关研究报告舞蹈家或体操运动员的灰质体积都是减小的。那么脑对舞蹈训练的适应到底是协调各脑区皮层以增减互补的方式实现?还是以减小优化的方式达成?其二, 如何理解舞蹈训练与音乐训练对皮层体积影响的相反趋势? 我们发现现代舞训练组与弦乐训练组均在 M1出现显著的灰质体积改变, 但这种改变趋势却是相反的。这一结果否说明皮层的体积增加直接与局部功能提升相关,而减小则可能提示是在一个系统内发生整体性优化的结果。以上问题均需要进一步的实证研究进行阐释。
段旭君. (2013).基于大尺度脑网络分析方法的脑可塑性研究(博士学位论文).电子科技大学, 成都.
蒋存梅. (2016).音乐心理学. 上海: 华东师范大学出版社.
马清. (2000).音乐理论与管弦乐基础. 北京: 北京大学出版社.
平心. (2004).舞蹈心理学.北京: 高等教育出版社.
吕艺生. (2003).舞蹈学导论.上海: 上海音乐出版社.
覃嫔. (2018).舞蹈艺术的训练研究.北京: 北京理工大学出版社.
周临舒, 赵怀阳, 蒋存梅. (2017). 音乐表演训练对神经可塑性的影响: 元分析研究.心理科学进展, 25(11), 1877–1887.
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry--the methods.NeuroImage, 11, 805–821.
Ashburner, J., & Friston, K. J. (2005). Unified segmentation.NeuroImage, 26(3), 839–851.
Bangert, M., Peschel, T., Schlaug, G., Rotte, M., Drescher, D.,Hinrichs, H., ... Altenmüller, E. (2006). Shared networks for auditory and motor processing in professional pianists:Evidence from fMRI conjunction.NeuroImage, 30(3),917–926.
Bangert, M., & Schlaug, G. (2006). Specialization of the specialized in features of external human brain morphology.European Journal of Neuroscience, 24(6), 1832–1834.
Baumann, S., Koeneke, S., Schmidt, C. F., Meyer, M., Lutz,K., & Jancke, L. (2007). A network for audio-motor coordination in skilled pianists and non-musicians.Brain Research, 1161, 65–78.
Bermudez, P., Lerch, J. P., Evans, A. C., & Zatorre, R. J.(2009). Neuroanatomical correlates of musicianship as revealed by cortical thickness and voxel-based morphometry.Cerebral Cortex, 19(7), 1583–1596.
Bostan, A. C., Dum, R. P., & Strick, P. L. (2013). Cerebellar networks with the cerebral cortex and basal ganglia.Trends in Cognitive Sciences, 17(5), 241–254.
Brown, S., Martinez, M. J., & Parsons, L. M. (2006). The neural basis of human dance.Cerebral C ortex, 16(8),1157–1167.
Burzynska, A. Z., Finc, K., Taylor, B. K., Knecht, A. M., &Kramer, A. F. (2017). The dancing brain: Structural and functional signatures of expert dance training.Frontiers in Human Neuroscience, 11, 566.
Calvo-Merino, B., Glaser, D. E., Grèzes, J., Passingham, R.E., & Haggard, P. (2005). Action observation and acquired motor skills: An FMRI study with expert dancers.Cerebral Cortex, 15(8), 1243–1249.
Cross, E. S., Hamilton, A. F., & Grafton, S. T. (2006). Building a motor simulation de novo: Observation of dance by dancers.NeuroImage, 31(3), 1257–1267.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn,U., & May, A. (2004). Neuroplasticity: Changes in grey matter induced by training.Nature, 427(6972), 311–312.
Giacosa, C., Karpati, F. J., Foster, N. E. V., Penhune, V. B.,& Hyde, K. L. (2016). Dance and music training have different effects on white matter diffusivity in sensorimotor pathways.NeuroImage, 135, 273–286.
Groussard, M., Rauchs, G., Landeau, B., Viader, F.,Desgranges, B., Eustache, F., & Platel, H. (2010). The neural substrates of musical memory revealed by fMRI and two semantic tasks.NeuroImage, 53(4), 1301–1309.
Groussard, M., Viader, F., Landeau, B., Desgranges, B.,Eustache, F., & Platel, H. (2014). The effects of musical practice on structural plasticity: The dynamics of grey matter changes.Brain and Cognition, 90, 174–180.
Han, Y., Yang, H., Lv, Y. T., Zhu, C. Z., He, Y., Tang, H.H., ... Dong, Q. (2009). Gray matter density and white matter integrity in pianists' brain: A combined structural and diffusion tensor MRI study.Neuroscience Letters, 459(1),3–6.
Hänggi, J., Koeneke, S., Bezzola, L., & Jäncke, L. (2010).Structural neuroplasticity in the sensorimotor network of professional female ballet dancers.Human Brain Mapping,31(8), 1196–1206.
Huang, H. Y., Wang, J. J., Seger, C., Min, L., Feng, D., Wu,X. Y., ... Huang, R. W. (2017). Long-term intensive gymnastic training induced changes in intra- and inter-network functional connectivity: An independent component analysis.Brain Structure and Function, 223(1), 131–144.
Huang, R. W., Lu, M., Song, Z., & Wang, J. (2015).Long-term intensive training induced brain structural changes in world class gymnasts.Brain S tructure and Function, 220(2), 625–644.
Hutchinson, S., Lee, L. H. L., Gaab, N., & Schlaug, G.(2003). Cerebellar volume of musicians.Cerebral Cortex,13(9), 943–949.
Hyde, K. L., Peretz, I., & Zatorre, R. J. (2008). Evidence for the role of the right auditory cortex in fine pitch resolution.Neuropsychologia, 46(2), 632–639.
Jola, C., McAleer, P., Grosbras, M. H., Love, S. A., Morison,G., & Pollick, F. E. (2013). Uni- and multisensory brain areas are synchronised across spectators when watching unedited dance recordings.i-Perception, 4(4), 265–284.
Jones, J., Adlam, A., Benattayallah, A., & Milton, F. (2017,July).Working memory training increases recruitment ofthe middle fr ontal g yrus in children. Poster session presented at the Conference of Experimental Psychology Society, Reading, UK.
Karpati, F. J., Giacosa, C., Foster, N. E. V., Penhune, V. B.,& Hyde, K. L. (2017). Dance and music share gray matter structural correlates.Brain Research, 1657, 62–73.
Kheradmand, A., & Zee, D. S. (2011). Cerebellum and ocular motor control.Frontiers in Neurology, 2, 53.
Koelsch, S., & Siebel, W. A. (2005). Towards a neural basis of music perception.Trends in Cognitive Sciences, 9(12),578–584.
Lahav, A., Saltzman, E., & Schlaug, G. (2007). Action representation of sound: Audiomotor recognition network while listening to newly acquired actions.Journal o f Neuroscience, 27(2), 308–314.
Laufer, I., Negishi, M., Lacadie, C. M., Papademetris, X., &Constable, R. T. (2011). Dissociation between the activity of the right middle frontal gyrus and the middle temporal gyrus in processing semantic priming.PLoS O ne, 6(8),e22368.
Li, G. J., He, H., Huang, M. T., Zhang, X. X., Lu, J., Lai, Y.X., ... Yao, D. Z. (2015). Identifying enhanced corticobasal ganglia loops associated with prolonged dance training.Scientific Reports, 5, 10271.
Li, S. Y., Han, Y., Wang, D. Y., Yang, H., Fan, Y. B., Lv, Y.T., ... He, Y. (2010). Mapping surface variability of the central sulcus in musicians.Cerebral Cortex, 20(1), 25–33.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D.,Ashburner, J., Frackowiak, R. S. J., & Frith, C. D. (2000)Navigation-related structural change in the hippocampi of taxi drivers.Proceedings of the National Academy ofSciencesof the United States of America, 97(8), 4398–4403.
Mutschler, I., Schulze-Bonhage, A., Glauche, V., Demandt,E., Speck, O., & Ball, T. (2007). A rapid sound-action association effect in human insular cortex.PLoS One, 2(2),e259.
Nichols, T. E. (2012). Multiple testing corrections,nonparametric methods, and random field theory.NeuroImage, 62(2), 811–815.
Oldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburgh inventory.Neuropsychologia,9(1), 97–113.
Ono, Y., Nomoto, Y., Tanaka, S., Sato, K., Shimada, S.,Tachibana, A., ... Noah, J. A. (2014). Frontotemporal oxyhemoglobin dynamics predict performance accuracy of dance simulation gameplay: Temporal characteristics of top-down and bottom-up cortical activities.NeuroImage,85, 461–470.
Öztürk, A. H., Tasçioglu, B., Aktekin, M., Kurtoglu, Z., &Erden, I. (2002). Morphometric comparison of the human corpus callosum in professional musicians and nonmusicians by using in vivo magnetic resonance imaging.Journal of Neuroradiology, 29(1), 29–34.
Rüber, T., Lindenberg, R., & Schlaug, G. (2015). Differential adaptation of descending motor tracts in musicians.Cerebral Cortex, 25(6), 1490–1498.
Schlaug, G., Jancke, L., Huang, Y., & Steinmetz, H. (1995).In vivo evidence of structural brain asymmetry in musicians.Science, 267(5198), 699–701.
Schneider, P., Scherg, M., Dosch, H. G., Specht, H. J.,Gutschalk, A., & Rupp, A. (2002). Morphology of Heschl's gyrus reflects enhanced activation in the auditory cortex of musicians.Nature Neuroscience, 5(7), 688–694.
Shibasaki, H., Sadato, N., Lyshkow, H., Yonekura, Y., Honda,M., Nagamine, T., ... Konishi, J. (1993). Both primary motor cortex and supplementary motor area play an important role in complex finger movement.Brain, 116,1387–1398.
Sluming, V., Barrick, T., Howard, M., Cezayirli, E., Mayes,A., & Roberts, N. (2002). Voxel-based morphometry reveals increased gray matter density in Broca's area in male symphony orchestra musicians.NeuroImage, 17(3),1613–1622.
Taubert, M., Draganski, B., Anwander, A., Muller, K.,Horstmann, A., Villringer, A., & Ragert, P. (2010). Dynamic properties of human brain structure: Learning-related changes in cortical areas and associated fiber connections.Journal of Neuroscience, 30(35), 11670–11677.
Turner, R. S., Grafton, S. T., Votaw, J. R., Delong, M. R., &Hoffman, J. M. (1998). Motor subcircuits mediating the control of movement velocity: A PET study.Journal of Neurophysiology, 80(4), 2162–2176.
