地盘松火后自然更新能力比较*
2018-04-25张辉红黄博强余婷孟丽媛曹建新张光飞苏文华
张辉红,黄博强,余婷,孟丽媛,曹建新,张光飞,苏文华
(1.云南大学 生态学与地植物学研究所,云南 昆明 650091; 2.云南省林业科学院,云南 昆明650201)
野火是一种世界广泛存在的现象,除了四季潮湿的森林以外,几乎所有的森林都曾在某一时期着过火[1]。在气候变化和人为干扰的影响下,过去几十年全球范围内森林火灾发生频率及火灾面积显著增加[2-4]。在易火地区,植物产生了适应火烧的对策,即通过萌生与实生2种途径进行更新恢复。植被对火干扰的响应及火烧后植被更新动态是目前火生态研究的核心问题之一,有助于人们理解火的作用和预测火烧后森林生态系统的结构功能变化[5-6]。云南是森林火高发区,森林防火、火烧后的群落重建和生态恢复已经成为该区域的研究热点和管理难点[7-8]。
地盘松(Pinusyunnanensisvar.pygmaea)是云南松的一个灌木状变种,常生长在土壤贫瘠干旱地段,是易发生火灾的群落之一[9]。火烧对地盘松原植株的地上部分危害严重,火后更新能力对其在易火生境中的生存极为重要。地盘松与云南松(P.yunnanensis)一样球果成熟后宿存在植冠形成植冠种子库[10]。火烧高温可诱导地盘松球果裂开并释放种子,还可促进其种子萌发。与云南松不同,地盘松还具有火后萌生能力,这一特性在松属植物中并不多见[9,11]。然而,目前对地盘松火烧后的自然更新情况尚不了解。本文通过调查昆明市西山区团结街道办事处棋台社区利者村后山发生火烧的地盘松林,研究比较了地盘松火烧后的自然更新恢复方式。研究结果对于了解地盘松对火烧的适应对策和火烧后的科学管理具有指导意义。
1 材料和方法
1.1 研究区自然概况
团结街道办事处位于云南省昆明市西山区西北部,地处102°20′58″-102°36′54″E,24°58′25″-25°12′51″N,总面积约为41 121hm2,其中林地面积约为34 000hm2。属于半湿润季风气候,境内具有四季分明、夏无酷暑、冬少严寒、干湿季分明等特点。年平均气温约为14℃,最高气温28℃,最低气温-5.4℃,日较差9-11℃。≥10℃有效积温2 600-5 000℃,无霜期约257d,降水量800-1 200mm。2014年4月16日1500时,昆明市西山区团结街道办事处棋台社区利者村后山发生森林火灾,过火面积约为210hm2。
1.2 野外调查方法
林火2年后,于2016年5月6-10日,对研究区域地盘松林火烧后更新恢复情况进行了详细调查。在火烧区域对3种不同立地条件(阳坡、阴坡、坡顶)分别设置了6个10m×10m的调查样方,每个样方面积为100m2,共计18个样方。对于每个样方,调查内容有火烧前地盘松原植株的株丛数、枝条数以及火烧后地盘松更新植株的株丛数、枝条数、株高和地径。由于过火区域内地盘松树干基本保存,且树与树之间也能明显区别,故火烧前原植株的数据可用过火后原植株的数据来表示。
1.3 数据处理与分析
利用Excel 2003软件对数据进行统计分析并作图,采用SPSS 22.0软件通过95%水平上进行单因素方差(ANOVA)分析,Duncan检验法进行多重比较及差异显著性检验(α=0.05)。
2 结果与分析
2.1 火烧前的状态及火烧后的受损情况
在样地火烧前,不同立地之间地盘松原植株的枝条数和株丛数存在显著差异。这可能是因为不同坡向的立地小气候不同。不同坡向的土层厚度、土壤水分和光照强度等条件均存在差异[12]。调查表明,阴坡、阳坡和坡顶上的枝条数分别为(37.40±7.47)枝/100m2、(59.60±14.54)枝/100m2、(152.50±13.77)枝/100m2,坡顶的枝条数最多,显著高于阴坡和阳坡的枝条数。阳坡的枝条数显著多于阴坡的枝条数。坡顶的株丛数也最多,达到了(30.25±3.59)丛/100m2,显著高于阴坡(16.20±1.30)丛/100m2和阳坡(22.00±5.34)丛/100m2,阳坡的株丛数显著多于阴坡的株丛数(图1)。

图1 火烧前不同坡向样地中地盘松枝条数和株丛数的比较
在所调查的18个样地中,受损比例为0-20%的样地数量最多,占所有样地的1/3;受损比例为20%-40%、80%-100%的样地数量最少,分别有2个;受损比例为40%-60%、60%-80%的样地数量均为4个。其中,受损比例大于40%的样地有10个,占全部的55.56%(图2)。这与地盘松生长在干旱贫瘠的特殊环境有关,当地盘松林地地表可燃物的载量达到较大值,在非常干燥的状况下,很容易被引燃。同时,地盘松林地地表可燃物具有很大的燃烧能量,且地盘松多为灌木状,一旦着火,常形成高强度的林冠火,难以扑救[13-14]。
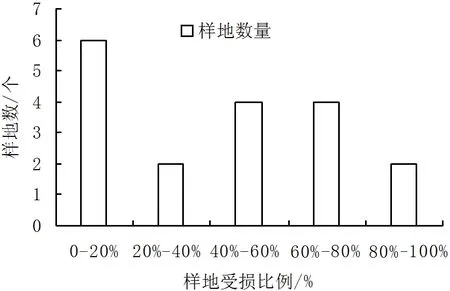
图2 火烧后样地中地盘松的受损比例
2.2 火烧后的地盘松恢复更新能力比较
地盘松群落地段过火2年后,过火地段仅地上部分死亡的原植株基本萌生出新枝条,还有种子萌发形成实生幼苗,种子更新幼苗已分支呈丛生状。更新株丛数在空间上有较大变化,阴坡、阳坡、坡顶的更新株丛数分别为(16.80±5.12)丛/100m2、(26.00±4.58)丛/100m2、(42.00±8.00)丛/100m2。坡顶的更新株丛数显著多于其原株丛数。阳坡和阴坡的更新株丛数分别多于各自的原株丛数,但差异不显著(P>0.05)。坡顶的更新株丛数显著多于阳坡和阴坡的更新株丛数。阳坡的更新株丛数大于阴坡的更新株丛数,但差异不显著。坡顶和阳坡的更新恢复能力较好,更新株丛数分别比原株丛数多38.84%和18.18%。阴坡的更新恢复能力相对较弱,更新株丛数仅比原株丛数多3.70%(图3)。
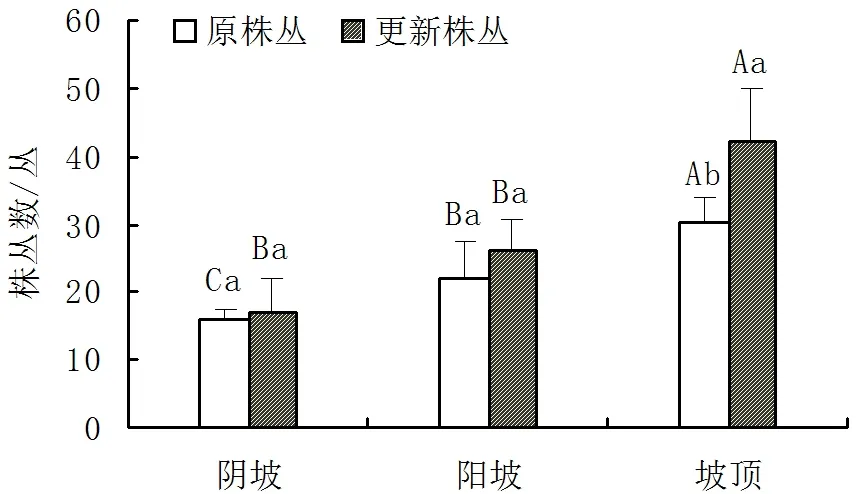
图3 火烧后不同坡向样地中地盘松更新株丛数与原株丛数的比较
更新植株的枝条数高于过火前的枝条数。阴坡、阳坡、坡顶的更新枝条数分别为(55.00±21.44)枝/100m2、(121.00±12.17)枝/100m2、(264.67±52.20)枝/100m2。坡顶和阳坡的更新枝条数分别显著多于各自的原枝条数。阴坡的更新枝条数多于其原枝条数,但差异不显著。坡顶的更新枝条数显著多于阳坡和阴坡的更新枝条数。阳坡的更新枝条数显著多于阴坡的更新枝条数。坡顶和阳坡的更新恢复能力较强,更新枝条数分别比原枝条数多73.55%和103.02%;阴坡的更新恢复能力相对较弱,更新枝条数比原枝条数多47.06%(图4)。

图4 火烧后不同坡向样地中地盘松更新枝条数与原枝条数的比较
2.3 火烧后2种更新途径的比较
在幼苗和幼树阶段,萌生枝的生长远远要比实生苗快[15]。究其原因,萌生枝可以利用原植株强大的根系以及树木其余部分所储存的丰富的代谢产物,并且占据较高的位置从而使之不易被遮光,因此其植株生长较快[16]。
样地火烧后,阴坡、阳坡和坡顶的地盘松更新恢复的萌生枝的高生长量分别为(62.22±10.64)cm、(54.97±10.24)cm和(49.28±4.25)cm,实生苗的高生长量阴坡为(29.98±11.78)cm、阳坡为(24.35±3.96)cm、坡顶为(25.00±0.00)cm。阴坡、阳坡和坡顶萌生枝的高生长量分别显著大于各自实生苗的高生长量。阴坡萌生枝和实生苗的高生长量分别大于阳坡和坡顶的高生长量,但差异不显著(P>0.05)(图5)。

图5 火烧后不同坡向样地中地盘松萌生枝与实生苗高生长量的比较
与实生相比,萌生具有抗干扰能力强、幼年期生长迅速等特点,因而它在植被演替的前期阶段发挥着明显的促进作用。而当干扰强度较低、种间竞争关系稳定时,物种更倾向于采取实生更新的策略[17]。
样地火烧后,阴坡、阳坡和坡顶更新恢复的萌生枝的地径分别为(1.20±0.60)cm、(0.95±0.55)cm和(1.63±0.64)cm,实生苗的地径阴坡为(0.77±0.51)cm、阳坡为(0.57±0.30)cm、坡顶为(0.80±0.61)cm。阴坡、阳坡和坡顶萌生枝的地径均显著大于各自实生苗的地径。坡顶萌生枝的地径显著大于阴坡和阳坡萌生枝的地径。坡顶实生苗的地径大于阴坡和阳坡实生苗的地径,但差异不显著(图6)。这可能是因为高强度的火烧过后,对地盘松幼苗生长干扰强度大,而萌生枝的抗干扰能力强于实生苗,且原植株强大的地下根系为萌生枝提供了生长所需营养物质,因而使得萌生枝的地径大于实生苗。

图6 火烧后不同坡向样地中地盘松萌生枝与实生苗地径的比较
3 结论与讨论
森林更新是森林群落演替和植被生态恢复的重要生态学过程。干扰假说认为,火、风、滑坡、雪崩、动物、砍伐等自然和人为干扰是驱动森林萌生更新的主要因素[18]。本文研究对象为地盘松,其树形呈灌木状,当发生火灾时,基本都是林冠火,植株地上部分极易被火烧死,地下部分仍然可能存活。
研究结果表明,火烧后地盘松通过萌生与实生更新很快进行自然恢复,并且更新植株的枝条数和株丛数均高于原植株。坡顶和阳坡的地盘松更新恢复能力较好,更新株丛数分别比原株丛数多38.84%和18.18%,更新枝条数分别比原枝条数多73.55%和103.02%;而阴坡的更新恢复能力相对较弱,更新株丛数比原株丛数多3.70%,更新枝条数比原枝条数多47.06%。已有研究表明,火后更新群落的物种多样性受到4个因素的影响,即地形和土壤环境、火烧强度、火前植被和物种生活史特征[19]。对于坡顶而言,可能地势相对平坦,火烧后土壤的灰分和养分不易流失,有利于地盘松幼苗的恢复和生长。一般来讲,坡度越陡,养分质量分数越低。这是因为坡度越大,土壤养分越容易流失[20]。火烧不久后,滇中迎来雨季,阳坡光照条件普遍好于阴坡,从而迅速长出大量幼苗。研究结果还表明,萌生枝的长势明显优于实生苗,阴坡、阳坡和坡顶更新恢复的萌生枝的高生长量均大于实生苗,萌生枝的地径也大于实生苗。因为萌生植株能通过其原有的强大根系,更有效地利用土壤中的养分资源,通常比实生植株生长快,具有更强的环境适应能力[21-22]。萌生更新在调节萌生和实生2种更新方式的平衡上具有重要意义[18],地盘松通过萌生和实生2种更新方式的互补来适应环境,以提高其自身在生境中的存活能力。
综上所述,火烧后地盘松具有较强的更新恢复能力,可通过萌生和实生2种更新方式进行植被恢复。火烧高温可以促进地盘松植冠种子库释放种子形成土壤种子库,最终形成实生苗。地盘松具有强大的地下根系,火烧后地上部分死亡,地下部分仍有存活,能很快萌生出新枝。因此,林业相关部门今后在对类似火烧后的群落进行更新恢复时,可以采取封山育林的方式,避免过多的人为干扰,让其自然更新。
参考文献:
[1]Bowman D M,Balch J K,Artaxo P,etal.Fire in the earth system[J].Science,2009,324(24):481-484.
[2]Roder A,Hill J, Duguy B,etal.Using long time series of Landsat data to monitor fire events and post-fire dynamics and identify driving factors.A case study in the Ayora region (eastern Spain)[J].Remote Sensing of Environment,2008,112(1):259-273.
[3]Bowman D M,Balch J K,Artaxo P,etal.The human dimension of fire regimes on earth[J].Journal of Biogeography,2011,38(12):2223-2236.
[4]Oliveira S,Pereira J,San-Miguel-Ayanz J,etal.Exploring the spatial patterns of fire density in Southern Europe using geographically geighted regression[J].Applied Geography,2014,51:143-157.
[5]Casady G M,Van L W,Marsh S E.Evaluating post-wildfire vegetation regeneration as a response to multiple environmental determinants[J].Environmental Modeling & Assessment,2010,15(5):295-307.
[6]Forrestel E J, Donoghue M J,Smith M D.Convergent phylogenetic and functional responses to altered fire regimes in mesic savanna grasslands of North America and South Africa[J].New Phytologist,2014,203(3):1000-1011.
[7]赵凤君,舒立福,田晓瑞,等.1957-2007年云南省森林火险变化[J].生态学杂志,2009,28(11):2333-2338.
[8]田晓瑞,舒立福,赵凤君,等.未来情景下西南地区森林火险变化[J].林业科学,2012,48(1):121-125.
[9]金振洲,彭鉴.云南松[M].昆明:云南科学技术出版社,2004.
[10]Su W H,Shi Z,Zhou R,etal.The role of fire in the Central Yunnan Plateau ecosystem,southwestern China[J].Forest Ecology and Management,2015,356:22-30.
[11]Pausas J G.Evolutionary fire ecology:lessons learned from pines[J].Trends in Plant Science,2015,20(5):318-324.
[12]聂莹莹,李新娥,王刚.阳坡-阴坡生境梯度上植物群落α多样性与β多样性的变化模式及与环境因子的关系[J].兰州大学学报(自然科学版),2010,46(6):73-79.
[13]王秋华,徐伟恒,李世友,等.地盘松林地表可燃物的燃烧火行为[J].消防科学与技术,2015,34(3):281-283.
[14]王秋华,徐伟恒,李伟,等.滇中地区地盘松林地表凋落物的潜在能量研究[J].林业调查规划,2014,39(6):16-20.
[15]Kujansuu J,荆涛,马万里,等.阔叶红松原生次生林和杂木次生林紫椴的生态学特征[J].北京林业大学学报,2003,25(1):113-115.
[16]Bellingham P J,Tanner V,Healey J R.Sprouting of trees in Jamaican montane forests after a hurricane[J].Journal of Ecology,1994,82(4):747-758.
[17]陈沐,曹敏,林露湘.木本植物萌生更新研究进展[J].生态学杂志,2007,26(7):1114-1118.
[18]朱万泽,王金锡,罗成荣,等.森林萌生更新研究进展[J].林业科学,2007,43(9):74-82.
[19]韩杰,应凌霄,李贵祥,等.云南松混交林火烧迹地更新早期草本层物种多样性的空间格局[J].植物生态学报,2016,40(3):200-211.
[20]Bond W J ,Midgley J J.Ecology of sprouting in woody plants:The persistence niche[J].Trends in Ecology & Evolution,2001,16(1):45-51.
[21]Kauffman J B.Survival by sprouting following fire in tropical forests of the eastern Amazon[J].Biotropica,1991,23(3):219-224.
[22]Vesk P A,Westoby M.Sprouting ability across diverse disturbances and vegetation types worldwide[J].Journal of Ecology,2004,92(2):310-320.
