黄芩苷对热应激小鼠卵巢氧化损伤的保护作用
2018-04-18周颖璐姜忠玲曹荣峰李华涛田文儒
周颖璐 , 高 琛 , 姜忠玲 , 王 新 , 丛 霞 , 曹荣峰 , 李华涛 , 田文儒
(青岛农业大学动物医学学院 , 山东 青岛 266109)
高温是造成动物生殖机能降低的重要环境因素,环境温度超过体温调节能力时,家畜体温升高,并常伴随着机体的氧化损伤。卵巢氧化应激影响卵母细胞成熟、卵子的发育,甚至造成卵泡闭锁、细胞凋亡等。氧化应激时,过多的ROS可激活多条信号通路,调控细胞对氧化应激的反应。核转录相关因子2(nuclear factor-erythroid 2-related factors 2,Nrf2)是细胞对抗外来化合物及氧化损伤最重要的核转录因子[1]。Nrf2/KEAP1系统是氧化应激的感受器[2],Nrf2/KEAP1信号途径失调将加重氧化应激损伤。黄芩苷是良好的抗氧化剂,但是黄芩苷对热应激诱导小鼠卵巢氧化损伤和细胞凋亡的保护研究尚未见报道。本研究主要集中于黄芩苷对热应激小鼠卵巢氧化损伤指标、Nrf2通路相关蛋白表达以及相关凋亡蛋白的表达揭示黄芩苷是否减少热应激诱导小鼠卵巢氧化损伤和细胞凋亡。
1 材料与方法
1.1 试验动物分组及热应激处理 6~7周龄雌性昆明系小鼠,体重25±0.5 g,按光照/黑暗=14 h/10 h控光,适应饲养1周后用于试验。分组与热应激按参考文献[3-4]方法处理。
1.2 样品采集和处理 热应激处理2 h后,断颈处死小鼠,剖腹取卵巢组织,一侧卵巢投入10%中性福尔马林中固定,做H.E.染色及免疫组化,另一侧冻存于-80 ℃冰箱中,用于制备组织匀浆以及提取总蛋白。
1.2.1 卵巢组织匀浆制备及蛋白浓度的测定 称重卵巢组织并按重量∶体积=1∶19加入生理盐水,制成5%组织匀浆,并在4 ℃条件下3 000 r/min离心10 min,用Bradford法测定其上清中组织蛋白浓度。
1.2.2 组织总蛋白提取 称重卵巢组织并按重量∶体积=1∶10比例加入RIPA裂解液。用电动组织匀浆研磨器研磨至充分裂解,然后以12 000 r/min 离心5 min,将上清液于-20 ℃保存备用。
1.3 卵巢切片组织学观察 将卵巢组织浸泡于体积分数为10%中性福尔马林中,24 h后,常规梯度酒精脱水,二甲苯处理后做石蜡包埋切片,切片厚度为4 μm,经H.E.染色后在普通光学显微镜下观察卵巢的组织学变化。
1.4 检测MDA和NO含量及SOD、GSH-Px和CAT活性 MDA、NO、SOD、GSH-Px和CAT分别用相应试剂盒(南京建成生物工程研究所)测定,操作过程严格按试剂盒说明书进行,依次加入相应量的卵巢组织匀浆和试剂,最后在紫外可见分光光度计上进行测定。
1.5 卵巢组织中Nrf2、Keap1、HO-1和NQO1检测 提取各组小鼠卵巢组织总蛋白,混合等体积2×SDS样品缓冲液,加热变性后进行SDS-PAGE凝胶电泳,在4 ℃条件下220 mA恒流湿转2 h,10% BSA室温封闭2 h,4 ℃条件下一抗(1∶1 000)孵育过夜,二抗(1∶3 000)室温孵育1 h,用Easy See Western Blot Kit试剂盒暗室内化学发光,最后用UVP凝胶成像系统拍照检测。
1.6 免疫组化法检测卵巢组织中Bcl-2、Bax、Caspase 3蛋白表达 对石蜡包埋的卵巢组织切片,二甲苯脱蜡,采用SABC法进行免疫组化检测,其操作步骤按SABC试剂盒的说明书进行。阴性对照组用PBS代替首抗。对于染色后的切片,在光学显微镜下观察,卵巢细胞胞质出现棕黄色颗粒为阳性结果。
1.7 数据分析 采用GraphPad Prism 5.01软件做统计学处理,组间差异用Dunnett单因素方差分析,均数间进行t检验,P<0.05有统计学意义。
2 结果与分析
2.1 小鼠卵巢组织H.E.染色变化 H.E.染色试验结果表明,常温对照组和黄芩苷组小鼠卵巢组织与卵泡细胞形态结构正常。热应激组小鼠卵巢组织组织细胞排列疏松,组织间质增宽,血管扩张并充满血液,组织部分细胞着染增深。与热应激组相比热应激加黄芩苷组卵巢组织,其各部分细胞形态正常,与对照组无明显差异(见中插彩版图1)。
2.2 卵巢组织内MDA和NO含量及SOD、GSH-Px和CAT活性的变化 与常温对照组相比,41 ℃热应激组中MDA(P<0.05)和NO(P<0.01)的含量均显著增加,SOD、GSH-Px和CAT活力均极显著降低(P<0.01)提示卵巢组织发生氧化应激反应。与41℃热应激组相比,热应激加黄芩苷组中MDA和NO含量显著降低(P<0.05),SOD(P<0.01)、GSH-Px(P<0.05)和CAT(P<0.05)活性则显著升高,抗氧化酶活性恢复。而热应激加黄芩苷组与常温对照组和黄芩苷组相比无统计学差异,提示黄芩苷能够抑制热应激所致卵巢组织的氧化损伤(见图2)。
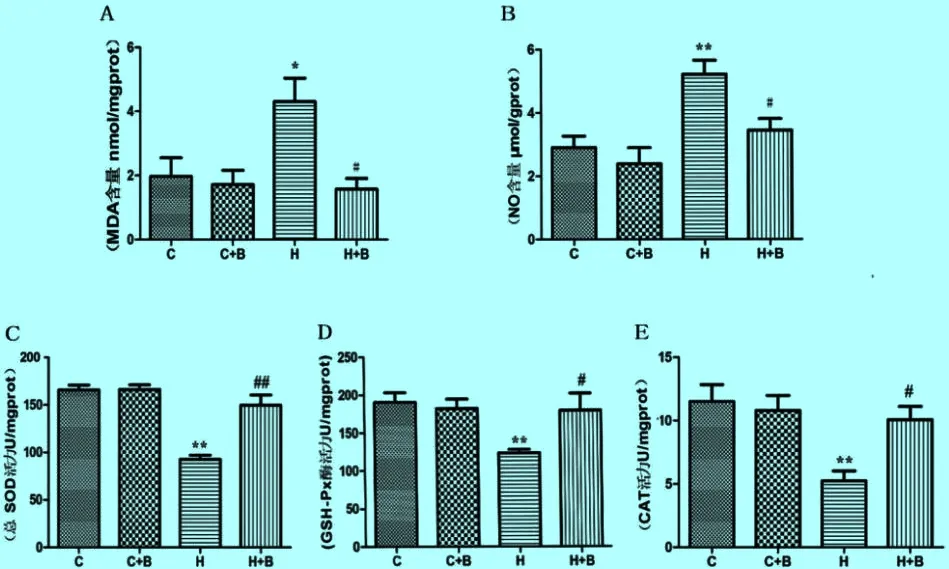
图2 黄芩苷对热应激小鼠卵巢组织内MDA(A)和NO(B)含量及T-SOD(C)、GSH-Px(D)和CAT(E)活性的影响
*:P<0.05(与常温对照组比差异显著); **:P<0.01(与常温对照组比差异极显著);#:P<0.05(与41 ℃热应激组比差异显著)
2.3 黄芩苷对热应激卵巢组织中Nrf2通路相关蛋白表达的影响
2.3.1 黄芩苷对Nrf2和Keap1蛋白表达的影响 结果显示,41 ℃热应激后小鼠卵巢组织中Nrf2和Keap1的表达显著增高(P<0.05)。而黄芩苷干预后Nrf2和Keap1表达显著降低,并与常温对照组和黄芩苷组相比无显著差异(见图3)。

图3 Nrf2(A)和Keap1(B)在卵巢组织内的表达
2.3.2 黄芩苷对HO-1和NQO1的表达的影响 结果显示,与对照组相比,41℃热应激后小鼠卵巢组织中HO-1和NQO1的表达均显著升高(P<0.05);而与41℃热应激组相比,热应激加黄芩苷组HO-1和NQO1的表达显著降低(P<0.05),并与常温对照组和黄芩苷组相比无显著差异(见图4)。

图4 各组小鼠卵巢组织内HO-1(A)和NQO1(B)蛋白的表达
2.4 免疫组化法检测黄芩苷对热应激卵巢组织中Bax、Bcl-2、Caspase 3蛋白表达的影响 免疫组织化学的方法检测Bax、Bcl-2和Caspase 3蛋白阳性反应主要定位于细胞膜或细胞质,染色呈淡黄色至棕黄色颗粒。与对照组相比,热应激能引起小鼠卵巢组织中Bax升高,特别是卵泡颗粒细胞棕黄色浓染;与热应激组相比,黄芩苷降低了热应激引起的卵泡颗粒细胞Bax表达。Bcl-2在正常卵巢组织中阳性表达,与对照组相比,热应激能降低卵泡颗粒细胞和间质细胞中Bcl-2的表达;与热应激组相比,黄芩苷能显著增加卵巢颗粒细胞与间质细胞中Bcl-2的表达,表现棕黄色浓染。Caspase 3在正常组织中呈现阴性表达,与对照组相比,卵巢间质细胞中Caspase 3深色黄染,呈现高表达,卵泡颗粒细胞中部分阳性表达;与热应激组相比,黄芩苷阻止热应激引起的Caspase 3表达,呈现阴性。
3 讨论
热应激使机体产生ROS,产生氧化损伤[5]。氧化损伤是许多外源化合物导致机体毒性作用的主要早期分子机制之一,生物体内ROS,包括NO等堆积是反应机体氧化应激状态的直接证据。MDA是细胞内产生的 ROS攻击细胞生物膜上的多不饱和脂肪酸,致使细胞生物膜破坏而形成的。MDA显著性增高提示机体氧平衡失调[6]。MDA高低可以间接反映机体细胞受自由基攻击的严重程度。本研究中,热应激小鼠卵巢组织细胞形态结构异常,出现充血、水肿等现象。小鼠遭受热应激后卵巢组织中NO及MDA的含量增加,抗氧化酶(SOD、GSH-Px、CAT)的活性降低,提示热应激诱导生成自由基进而降低动物抗氧化能力。李忠浩等研究发现,奶牛外周血液中SOD、GSH-Px的活性在夏季高温季节极显著下降,MDA含量极显著上升[7]。热应激下公兔睾丸内的SOD、CAT水平显著下降[8],即睾丸组织中抗氧化酶活性降低,导致清除自由基的能力下降,自由基生成过多,睾丸组织细胞受损[9]。因此我们推断小鼠抗氧化能力降低的同时氧化产物增多,小鼠卵巢发生氧化损伤。
在正常生理平衡下,Nrf2通过与 KEAP1相互作用并被蛋白酶降解而保存于细胞质内。当氧化应激作用下,Nrf2/KEAP1之间的反应被扰乱,导致Nrf2释放并入核,随后与MAF蛋白结合形成异二聚体,并与ARE结合,指导形成解毒酶、抗氧化蛋白及转录蛋白,从而起到细胞保护的作用[10]。在本研究中,小鼠遭受热应激后其卵巢组织中Nrf2的表达明显增高,Keap1的含量也有所升高。分析其原因,热应激导致小鼠卵巢组织中促氧化物质产生增多,刺激Nrf2表达上调,从而发挥其细胞保护作用,为了结合表达增多的Nrf2,组织细胞同时增加了Keap1的表达。同时,HO-1和NQO1的表达也明显上调,可能是Nrf2通过与ARE及bZIP蛋白结合后启动Ⅱ相解毒酶的表达。HO-1可以帮助神经元抵抗氧化应激损伤[11]。NQO1可增强C/EBPα 抵御20 S蛋白酶降解的稳定性,有效抵抗辐射对骨髓的氧化损伤[12]。ARE相关基因的表达上调使机体能够适应内环境ROS的积聚,证实KEAP1/Nrf2解聚可防御氧化应激损伤。而通过激活 Nrf1/Nrf2途径,可有效抑制 ROS的产生,以维持肾脏的功能[13]。双酚A通过反式激活异型的Nrf1/Nrf2途径,增加ARE靶基因HO-1、NQO1的表达,从而大大减少了人胚肾293细胞内环境的ROS,起到抗氧化应激的作用[14]。
国内外研究发现,细胞受到氧化应激会诱导产生凋亡,H2O2处理鼠胸腺细胞可诱导细胞发生明显的凋亡[15]。Bcl-2家族和Caspase家族在细胞凋亡转导通路尤其是线粒体途径中发挥了极其重要的调控作用。本研究也发现,热应激能在一定程度上明显的上调Bax,下调Bcl-2和Caspase 3蛋白水平。 Bax/Bcl-2比值可直接导致线粒体膜的功能紊乱,导致线粒体膜电位的丢失,以及促凋亡蛋白的释放。Caspase与线粒体通路、死亡受体通路以及内质网通路紧密相关。其中,Caspase与线粒体介导的凋亡密切相关。当线粒体受到凋亡信号的刺激,其释放的Cyt-C能与Apaf-1及Caspase-9的复合物形成凋亡体,导致Caspase-9的自动活化 ,从而引发级联反应,激活下游的Caspase 3,最终导致凋亡。
黄芩苷对乙酰氨基酚和四氯化碳引起的肝脂质过氧化损伤以及对铁蛋白(或柠檬酸铁)-亚硝酸盐-过氧化氢体系造成的肝氧化损伤均有保护作用[16]。黄芩苷可以抑制热应激下LLC-PK1凋亡,其作用机制可能与影响凋亡基因Bax和Bcl-2表达有关[17],这与研究三黄泻心汤的主要成分黄芩苷对氧化损伤的保护作用机制相同[18]。本研究发现,黄芩苷干预后小鼠卵巢组织其各部分细胞形态正常,未出现明显的病变。黄芩苷能够抑制MDA和NO的生成,提高抗氧化酶浓度,反映黄芩苷干预后小鼠卵巢组织细胞受ROS损伤的程度降低,黄芩苷对氧化产物造成的卵巢组织损伤有保护作用。黄芩苷可以有效降低Bax和Caspase 3蛋白在卵巢细胞中的表达,上调Bcl-2蛋白表达水平,抑制卵巢细胞的凋亡。同时Nrf2、Keap1、HO1和NQO1的表达均下降,可能由于在黄芩苷的干预,机体氧化还原平衡逐渐恢复,Nrf2转回入胞浆并通过泛素化降解,或者通过负反馈调节,促进Nrf2的消耗,Keap1、HO1和NQO1的消耗也同步增加,从而使抗氧化酶活性升高,清除自由基的能力增强,减少氧化产物的生成,缓解氧化应激带来的卵巢组织细胞凋亡。黄芩苷一方面通过其本身捕捉氧自由基,清除活性氧等氧化产物的能力,另一方面通过促进抗氧化酶的生成和Nrf2等蛋白的消耗来间接地使氧化产物浓度降低,保护组织不受损伤。因此可以推测,Nrf2/KEAP1信号通路可能参与黄芩苷抗氧化的过程。
参考文献:
[1] Park S Y, Choi S M, Park S J,etal. The role of nuclear factor-E2-related factor 1 in the oxidative stress response in osteoblast cells[J]. Bone, 2011, 48(48):S121-S121.
[2] Baird L,Dinkova-Kostova A T. The cytoprotective role of the Keap1-Nrf2 pathway[J]. Arch Toxicol, 2011,85 (4):241-272.
[3] 高琛,隋君霞,周颖璐,等. 黄芩苷对热应激小鼠睾丸氧化损伤的保护作用[J]. 农业生物技术学报,2017,25(09):1 470-1 477.
[4] Xia Cong, Qian Zhang, Huatao Li,etal. Puerarin ameliorates heat stress-induced oxidative damage and apoptosis in Sertoli cells through suppressing ROS production and upregulating Hsp72 expression[J].Theriogenology, 2017 Jan (15) 88:215-227.
[5] Valls-Belles V, Torres Mdel C, Boix L. Alpha-Tocopherol, MDA-HNE and 8-OHd G levels in liver and heart mitochondria of adriamycin-treated rats fed with alcohol-free beer[J].Toxicology,2008, 249:97-101.
[6] 李忠浩,孔丽娟,刘庆华,等. 不同温度下荷斯坦奶牛外周血抗氧化指标的变化及其与淋巴细胞凋亡的关系[J].中国奶牛,2007,29(S1):136-139.
[7] 彭海龙,杨淑华,何剑斌.热应激对兔睾丸和附睾CAT与SOD水平的影响[J].动物医学进展,2010,31(6):85-89.
[8] 梁明振,黄桂花,何烽杰,等.热应激对公兔精液品质、睾丸组织形态及抗氧化指标的影响[J]. 广西农业科学,2010,41(3):263-265.
[9] Sarkar S, Mukherjee S, Chattopadhyay A,etal. Low dose of arsenic trioxide triggers oxidative stress in zebrafish brain: Expression of antioxidant genes[J]. Ecotoxicology & Environmental Safety, 2014, 107(9):1-8.
[10] Chen J, Tu Y, Connolly E C,etal. Heme oxygenase-2 protects against glutathione depletion-induced neuronal apoptosis mediated by bilirubin and cyclic GMP[J]. Current Neurovascular Research, 2005, 2(2):121-131.
[11] Xu J, Jaiswal A K. NAD (P) H: quinone oxidoreductase 1 (NQO1) competes with 20S proteasome for binding with C/EBPα leading to its stabilization and protection against radiation-induced myeloproliferative disease[J].J Biol Chem,2012 Dec 7;287 (50):41 608-41 618.
[12] Saito H. Toxico-pharmacological perspective of the Nrf2-Keap1 defense system against oxidative stress in kidney diseases[J]. Biochemical Pharmacology, 2013, 85(7):865.
[13] Chepelev N L, Enikanolaiye M I, Chepelev L L,etal. Bisphenol A Activates the Nrf1/2-Antioxidant Response Element Pathway in HEK 293 Cells[J]. Chemical Research in Toxicology, 2013, 26 (3):498-506.
[14] 蒋定文, 郭明秋, 陈丽茵, 等. HSP70与热应激诱导胸腺细胞凋亡关系的研究[J]. 海军医学杂志, 2000(04): 303-306.
[15] 高中洪,黄开勋,徐辉碧. 黄芩黄酮对H2O2导致的神经细胞损伤的保护作用[J]. 中国药理学通报,2000(05): 589-590.
[16] 孙春玲, 国晓瞳, 赵园, 等. 黄芩苷对热应激条件下猪近端肾小管(LLC-PK1)细胞凋亡率及B细胞淋巴瘤-2基因(Bcl-2)和Bcl-2相关X蛋白基因(Bax)表达的影响[J]. 农业生物技术学报, 2014,22(12): 1 553-1 560.
[17] Liou S F, Hsu J H, Liang J C,etal. San-Huang-Xie-Xin-Tang protects cardiomyocytes against hypoxia/reoxygenation injury via inhibition of oxidative stress-induced apoptosis[J]. J Nat Med, 2012, 66(2): 311-320.
