高寒草甸土壤贮水量对封育措施的响应
2018-04-11贺慧丹祝景彬未亚西李红琴杨永胜李英年
贺慧丹, 祝景彬, 未亚西, 李红琴, 杨永胜, 李英年
(1.中国科学院 西北高原生物研究所, 西宁 810001; 2.中国科学院大学, 北京 100049;3.中国科学院 高原生物适应与进化重点实验室, 西宁 810001)
高寒矮嵩草(Kobresiahumilis)草甸广布于青藏高原及其周围山地,约占青藏高原可利用草场的25%[1]。由于高寒草甸植物具有“高蛋白、高脂肪、高热值、低纤维”三高一低的特点,牧草适口性好,是畜牧业发展的物质基础。近年来,随着气候变暖、人口急剧增长及家畜数量不断增加,高寒草甸地区草地逐步退化、水土流失加剧,导致该区草地生产力、土壤固碳持水能力不断下降[2-3]。正因如此,国务院划分不同生态功能区进行生态恢复与治理工作。2005年开始的《青海三江源自然保护区生态保护和建设总体规划》(生态治理工程)一期工程,经过几年的实施,三江源生态系统退化趋势得到初步遏制,重点生态建设工程区生态状况逐渐好转,但生态建设任务的长期性、艰巨性凸显[4]。
三江源生态治理工程中,封育禁牧是有效恢复和重建植被的办法之一。随封育时间延长,植被在正向演替的恢复过程中,其固碳持水能力、土壤水贮存量以及物种多样性均可发生改变[5-7]。然而,对于生态治理工程实施后生态环境的变化,以往的研究多集中在土壤有机碳、多样性、生物量[8]等方面。也有一些学者,如徐翠等[9]提出退化使土壤孔隙度等物理性质发生改变,从而影响土壤水分涵养能力。周印东等[10]认为自然植被的正向演替对表层土壤有机质含量有明显的促进作用,表层土壤田间持水量、容重、总孔隙度等与土壤持水性能相关的指标都与有机质含量呈显著或极显著相关。但较少涉及土壤贮水量、持水能力及与环境因素之间关系的研究。本研究以三江源玛沁县高寒草甸为研究区域,针对土壤贮水量对封育措施响应的问题,选择封育和自然放牧地为试验区,在定期测定土壤湿度、持水量、容重、孔隙度等指标的基础上,分析植被恢复过程中封育措施下土壤容重、持水能力、贮水量时空变化特征及环境影响机制,以期为生态治理工程实施下高寒草甸生态系统物质循环和能量流动的研究提供科学依据。
1 材料与方法
1.1 研究区概况
本研究在青海省果洛州玛沁县进行,玛沁地区位于青海省东南部,是国家级“三江源”重点生态保护区。玛沁县年平均气温-0.5℃,气温低,日温差大。年平均降水量514 mm,多集中在6—9月份,属高原大陆性半湿润气候,冬半年干燥少雨,夏半年凉爽湿润。植被为高寒草甸类型,优势种有矮嵩草、垂穗披碱草(Elymusnutans)、青藏苔草(CarexsiderostictaHance.)、异针茅(StipaalienaKeng)、高原早熟禾(PoaalpigenaLindm.)。伴生种有鹅绒委陵菜(Potentillaanserina)、异叶米口袋(GueldenstaedtiadiversifoliaMaxim.)、麻花艽(GentianastramineaMaxim.)、美丽风毛菊(Saussureasuperba)、黄花棘豆(Oxytropisochrocephala)、细叶亚菊(Ajaniatenuifolia)、矮火绒草(Leontopodiumnanum)、高原毛茛(Ranunculaceae)等[11]。植被群落结构简单、分异不明显。土壤为高山草甸土,土壤发育年轻,有机质含量高,粗骨性强。
1.2 研究材料与方法
1.2.1试验设计试验区设在玛沁县西北5 km处的果洛州气象局生态监测点100°12′E,34°28′N,海拔3 761 m的高寒草甸植被类型。封育样地系生态监测点于2003年封育,仅在春季的2—4月进行适度放牧,时长3个月。未封育样地为当地牧民自然放牧区,每年10月—翌年5月放牧,时长8个月。放牧强度均约1.3只羊单位/hm2。
观测区设在封育与未封育样地,在每个样地设计10 m×20 m的观测区,以中央点及10 m×20 m的角点附近为观测样点,即每个试验区共5个点(重复)。
1.2.2监测指标及方法土壤湿度及贮水量:供试分析的土壤水分监测于2013年5—9月每月8日,18日,28日在设计的上述5个重复点右侧4 m×4 m的范围内随机取样,每次分别用土钻收集0—10,10—20,20—30,30—40 cm层次土样装入铝盒,及时称取鲜重,带回实验室烘干至恒重后称干重,并计算其土壤质量含水率(占干土重的百分比)。土壤贮水量是指自然状况下一定土层厚度的土壤能够容纳的实际含水量,以土层深度的mm水量表示。有[12]:
W=F×H×SD×/10
(1)
式中:W为土壤贮水量(mm);F为土壤容重(g/cm3);H为土壤厚度(cm);SD为实测的土壤质量含水率;10代表把水层定为mm水量的转换系数。
植被生物量:于2013年8月底在设计的上述5个重复点左侧1 m处进行了50 cm×50 cm样方的生物量等植被群落特征调查。观测时先收集枯落物装袋,其次用剪刀齐地面分种剪下绿色植物装袋,并收集碎屑物,再用内径8 cm的根钻分0—10,10—20,20—40 cm层次收集土柱(每个样方3个重复,并同层次混合),土柱现场捣碎混合后过筛,将过筛后的土壤和带根系的土壤分别分层装袋。带根系的土壤带回实验室后分拣石粒、过筛、清洗等过程挑拣出包括多年死根在内的生物量装袋。在室内将地上生物量(绿体、枯落物)、碎屑、地下根系生物量等一并置入烘箱,在65℃烘温条件下烘干至恒重称重。
土壤容重、持水量:上述监测生物量的样方附近挖掘剖面,用内径为5 cm,容积为100 cm3的环刀分0—10,10—20,20—40 cm层次收集环刀土壤,取样后环刀迅速封盖并称量鲜重(m1),用透明胶带密封后带回实验室,室内清除胶带及污垢,揭去上底盖,仅留带网眼的下底盖放入平底容器中,注入水直至高度恰好在环刀上沿,在水中充分浸透12 h后,盖上底盖水平取出称重(m2);之后,把仅带有网眼底盖的环刀放在铺有干沙的平底盘中2 h,盖上底盖称重(m3);接着,把环刀放在平地盘上2昼夜,再称重(m4);最后在实验室烘箱烘干24 h(至恒重)后用电子天平(0.001 g)称重(m5)。计算前后差值并取平均值,测定得到土壤容重(g/cm3)、土壤持水量、毛管孔隙度(%)等。
降水量:采用果洛州气象局地面气象观测资料。
土壤有机质和有机碳密度:过筛后收集的土壤样品带回室内自然阴干,利用重铬酸钾容量法—外加热法测定土壤有机质的含量。有机碳密度(SOC,Kg C/m2)采用以下公式[13]:
(2)
式中:Di,Bi,OMi和S分别为土层厚度(cm)、土壤容重(g/cm3)、土壤有机碳含量(%)和对应面积(cm2);i代表土壤的分层数(i=1,2,3)。
1.2.3数据分析采用Excel 2013进行数据的基本处理和作图,SPSS 19.0软件的One-Way ANOVA统计分析指标的平均值、标准差及在p=0.05水平上各处理间的差异显著性。
2 结果与分析
2.1 土壤容重与土壤持水能力
图1给出了玛沁高寒草甸封育与未封育措施下土壤容重变化。由图1可知,封育与未封育土壤容重均随着深度增加而增加,最大可达1.27~1.39 g/cm3。
封育使不同层次的土壤容重均有所下降,与未封育区相比,0—40 cm整层土壤平均容重降低了11%,其中0—10 cm和10—20 cm差异达显著性差异(p<0.05),而20—40 cm无显著差异。
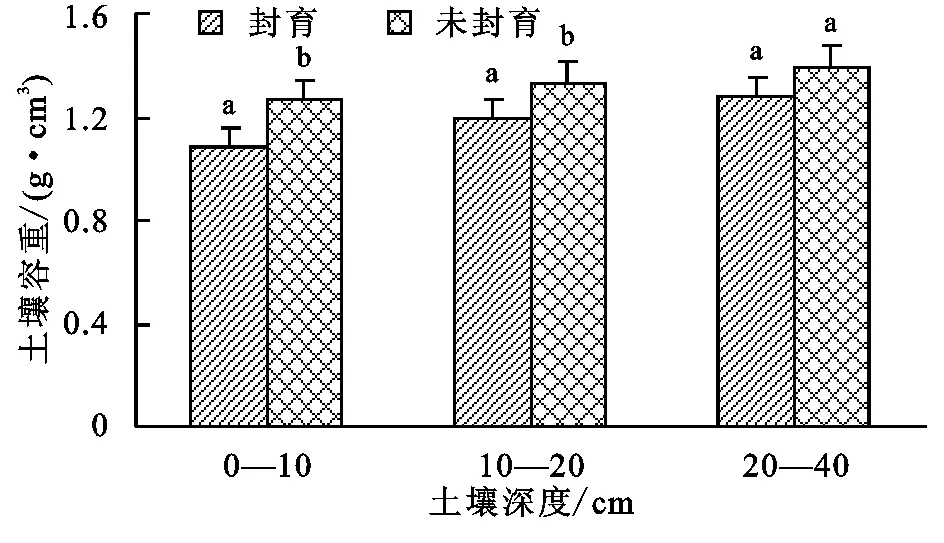
注:图中字母不同表示两者达到0.05显著水平,字母相同表示两者未达到显著水平。
图1封育和未封育土壤容重与土层深度的关系变化
封育11 a后土壤不同层次的最大持水力、毛管持水力、最小持水量均有所增加(表1),0—40 cm整层分别达201.96,177.37,156.61 mm,比未封育区分别高11.34,15.05,15.14 mm,分别增加了6%,9%和11%。封育11 a来,高寒草甸土壤饱和持水量、毛管持水量和田间持水量分别按1.03,1.37,1.38 mm/a的速率增加,封育11 a后毛管持水量和田间持水量增加速率较高。尽管封育样地与未封育样地持水能力差异均未达到显著水平(p>0.05),但封育后3个指标均高于未封育样地,说明封育措施能提高土壤的持水能力。
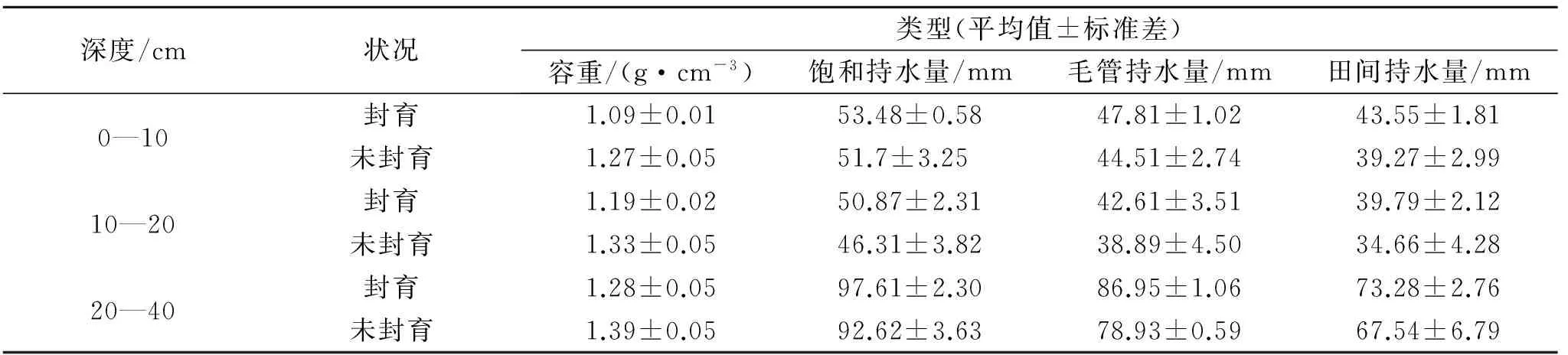
表1 土壤容重及土壤持水量
2.2 植物生长期土壤贮水量季节动态及与封育与非封育环境因素的关系
图2给出了2013年果洛玛沁高寒草甸封育与未封育措施下0—40 cm土壤层次贮水量以及降水量在植物生长期的季节动态。图2看到,封育与未封育措施下0—40 cm土壤层次贮水量动态变化基本一致,封育区0—40 cm层次土壤贮水量波动程度大于未封育样地且量值较高。两者贮水量差距在5月底6月初、8月底以后较为明显。在植物生长期的5—9月,封育与未封育措施下5月初到7月中旬土壤贮水量保持较高的水平,7月下旬以后,土壤贮水量下降明显,进入9月略有升高。土壤贮水量的这种变化不仅与降雨具有极大的相关性,而且与植物生长对土壤水的利用消耗有关。
从季节动态来看,5月到6月初,受低温环境影响,植物生长缓慢,叶面积较小,蒸腾作用不强,土壤蒸发也受到温度影响较弱,底层土壤(一般在40 cm层次以下)仍维持冻结状态,冻结层在融化过程中不断产生融冻水来补给土壤上层,同时冻结层的依然存在阻止了水分下渗,加之自然降水产生,促使了土壤贮水量增加。6月中旬到7月下旬,降水明显增多,但植被生长达到一定高度,叶面积处在增加阶段,有降水补给但也有较高的蒸散,二者共同作用下土壤贮水量也较高。其中,7月底封育区降水量具有最高值,但属于雨热同期,蒸发强烈,温度的升高和降雨的增加,植被快速生长,植被生长旺盛消耗大,这也是这一时期降水量大,但贮水量最高值却小于5月底的原因之一。也说明水分蒸散是高寒草甸地区降水损失的重要因素,蒸散降低相应的土壤贮水量就提高。2013年8月与多年平均降水量相比是偏少较多的一年,整个8月降水量仅为46.8 mm,是多年平均降水量的9.1%,降水偏少明显,加之植被蒸散在相对较高的温度环境下蒸散加大,导致8月土壤水散失严重,贮水量下降明显。进入9月后,随温度下降,日最低气温常处在0℃以下,植被生长减缓,叶面积处在下降阶段,植物进入枯黄期,地表面形成一定的枯落物,植物对其下面的地表产生最大的郁闭度,植被蒸腾受植物基本停止生长影响下降明显,土壤蒸发受较高的郁闭度影响而降低,土壤水分散失减少,加之温度低,土壤表面日消夜冻,使土壤水分容易积聚,缓慢提高了土壤贮水量。形成了土壤贮水量5月到6月初的融冻集聚期、6月中旬至9月初的波动消耗期和9月以后的蓄积期。
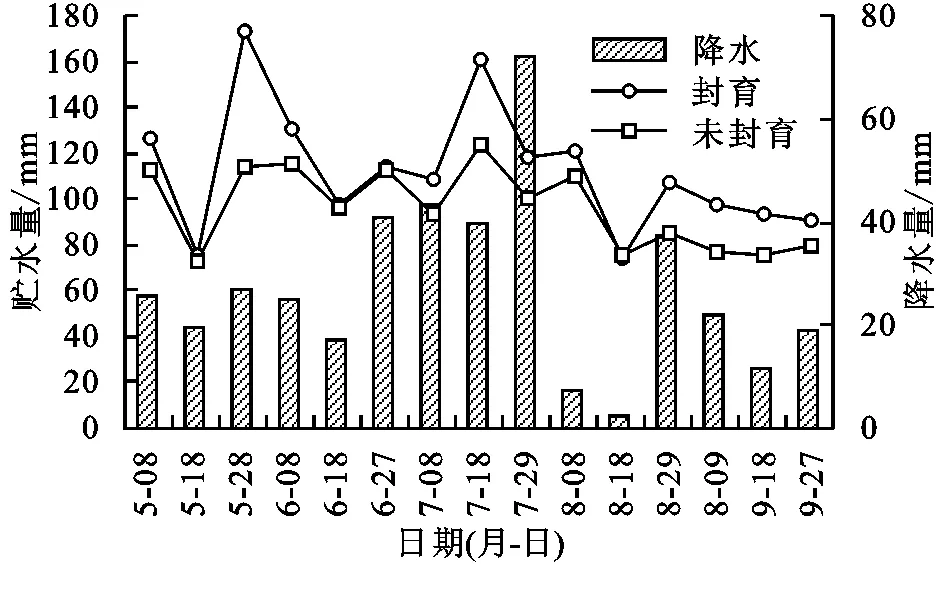
图2封育和未封育样地土壤贮水量及时段降水量的季节动态变化
统计发现,5—9月封育与未封育措施下0—40 cm层次土壤平均贮水量在73.38~173.23 mm范围波动,平均分别为112.58,96.26 mm。封育11 a后,土壤贮水量比未封育试验区高16.32 mm,平均按1.25 mm/a的速率增加。所表现的贮水量变化说明封育措施能有效提高土壤贮水量,利于土壤水源涵养。
分析生长期5—9月0—40 cm层次土壤平均贮水量与环境要素关系发现(表2,图3),5—9月平均土壤贮水量与土壤容重具有极显著负相关(图3A),与地下生物量、地上生物量和有机碳密度分别呈极显著正相关(图3B)、显著正相关(图3C)和无相关关系(图3D)。与土壤容重呈极显著负相关,说明随着土壤容重增加,土壤结实,孔隙度降低,大部分水分在地表被蒸发,土壤水分渗水率降低,土壤贮水率下降[14]。与地下生物量呈极显著正相关反映了植被虽然因植被生长好的状况下蒸腾提高,但较高的根系量是水分贮存的重要物质条件[15],同时,高寒草甸区的植被大多根系位于0—20 cm,尤其根系死后被分解留有的空隙,增加了土壤颗粒之间的孔隙度,水分下渗较多,贮水量高。地上生物量大,虽然对地面的覆盖可以减少土壤蒸发,但植被本身蒸腾作用较强,耗水明显,进而降低了土壤贮水量,由于地上生物量高低与植被覆盖度关系较大,绿体植物覆盖度增加可减少土壤水的蒸发,也可减少降水对土壤水的补给,但可增加植被的蒸腾,故实际上土壤贮水量与地上生物量的关系显得复杂了[16]。草地生态系统中有机碳主要储存于土壤中,其中土壤有机碳密度的大小取决于土壤容重、土壤有机质含量及土层厚度,容重增加,有机碳密度降低。封育样地未被啃食,枯落物较多,地面蒸发降低,而分解能使土壤有机质增加,提高有机碳密度,改善土壤结构,有利于土壤持水,贮水量增加。
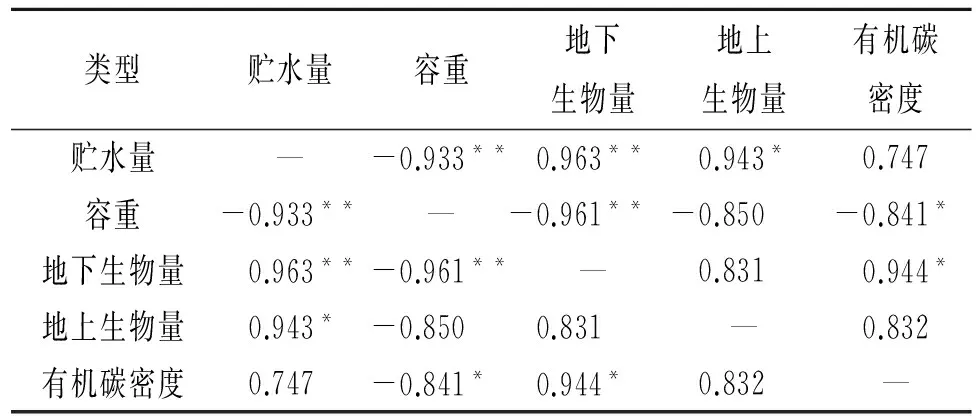
表2 土壤贮水量与容重、地下生物量、地上生物量以及有机碳密度的相关性检验
注:*表示显著相关性检验p<0.05;**表示极显著相关性检验p<0.01。
2.3 土壤贮水量的垂直分布
图4给出了封育与自然放牧的未封育样地0—40 cm层次土壤贮水量的垂直变化。可以看出,封育和未封育土壤贮水量在植物返青期(5月18日)和地上生物量达最大时期(8月18日)的上层0—10 cm和深层30—40 cm相反,而在强度生长期的7月18日和植物枯黄初期(9月18日),0—40 cm均表现为封育大于未封育。
图4A看到,在植物生长初期的5月18日,封育和未封育措施下,土壤20—30 cm深度土层贮水量均高于上层和下层,这与上层地面裸露,土壤已融化,底层仍有强度不大的土壤冻结有关。植物生长初期地表蒸发和较小的植被蒸腾导致上层土壤水散失相对较高而导致土壤贮水量低,而下层土壤仍维持一定的冻结层,其土壤水由冻结水和非冻结水组成,极大多非冻结水已通过土壤层次的温度梯度的作用补充到上一层次,在提高上层土壤贮水量的同时降低了下层土壤贮水量。
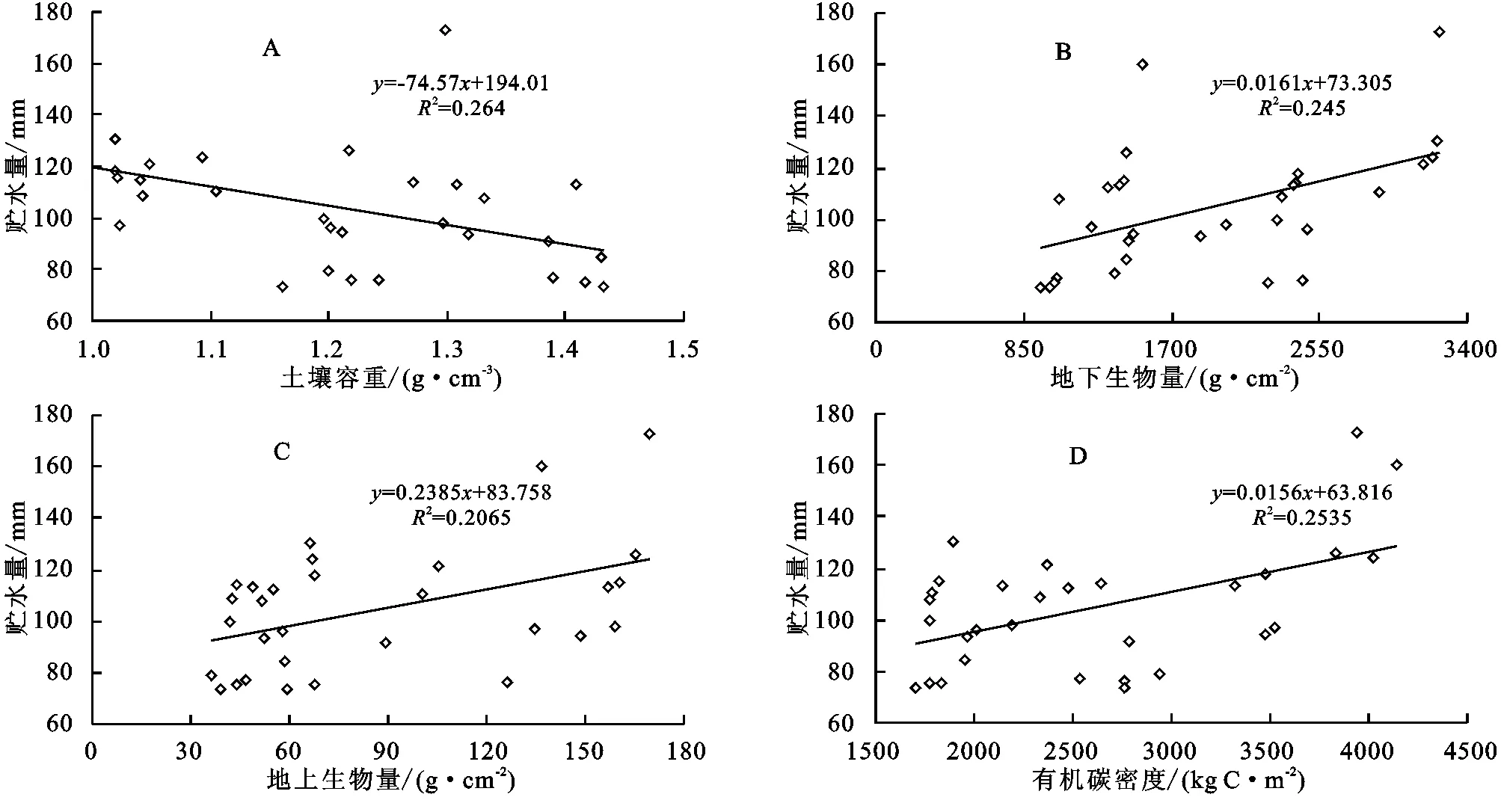
图3 封育与未封育样地5-9月平均土壤贮水量与土壤容重、地下生物量、地上生物量以及有机碳密度的关系
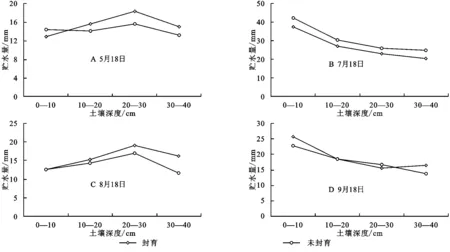
图4 封育与未封育样地土壤贮水量的垂直变化
植物进入强度生长期的7月18日(图4B),土壤贮水量在垂直方向上表现为随土层深度增加而降低。封育与未封育试验区垂直变化一致,在任何层次均表现为封育区明显高于未封育试验区,且因前期较高降水的影响,垂直变化上均表现出土壤表层高而深层低。7月是年内雨量最为丰沛时期,约达155.5 mm,占植被生长期5—9月降水量409.1 mm的38%,7月9日到7月18日期间,阶段降水量仍达40 mm。7月也是年内温度最高时期,降水量的增加及良好的热量条件下,植被加快生长,虽然植被蒸腾强,但不及降水的补给大,导致土壤上层土壤贮水量较高。由于植被根系大多分布在较浅,一定程度上阻止降水入渗土壤后的重力水下渗,使大部分水分集中于土壤上层,相对保持了深层土壤贮水量较低。
8月是植被地上生物量达最大时期,8月9日到8月18日(图4C)期间降水最少,仅为2.3 mm,而降水量的多少是影响土壤水分的主要因素[17]。这一时期温度较高,太阳辐射强,植被蒸腾和土壤蒸散强烈,丰富的地上生物量对降水有截流作用,使原本很少的降水很难渗入土壤,土壤水分散失量大,导致土壤上层贮水量达年内最低,而深层水由于重力作用得以保存,所以深层土壤贮水量高于上层土壤贮水量。
9月18日(图4D)植被基本进入枯落期,变化规律与7月18日相似,封育区与未封育区土壤表层贮水量分别为27.2,25.8 mm,相互之间差异很小,但受8月15日前后稍大降水量的影响,土壤水分的滞后效应下0—10 cm土层贮水量有所提高,且封育地大于未封育区。30—40 cm层次土壤贮水量差异逐渐减小。这时期植被生长逐渐停止,植被蒸散作用减弱,地表由于被枯落物和腐殖质覆盖蒸散作用也较弱,对降水的截流以及拦蓄使水分多集中于地表,这一时期地下生物量较高,能很好的涵养水分,故土壤浅层0—10 cm贮水量高于10—40 cm的深层贮水量。
3 讨论与结论
3.1 土壤容重与持水能力
张社奇等[18]的研究表明土壤容重会随着深度增加而增加,这与本研究的结论一致。土壤容重的大小主要受土壤孔隙度的影响,土壤疏松多孔,则容重越小,毛管吸水力越大,最大持水能力越大。通常来讲,土壤的最大持水能力越强,贮水能力越强,对水源的涵养功能越高。植物也通过提高土壤有机质含量改变土壤的结构,使土壤容重降低,并且使毛管孔隙度增大,有利于土壤的持水性[19-20]。本研究中放牧遭受家畜踩踏严重,土壤板结,土壤孔隙度减小,容重增加。而土壤持水能力与容重关系密切[21],放牧使土壤容重增加,渗透阻力加大,土壤的保水和持水能力下降。禁牧避免了家畜的影响,使土质变得松软,表层受到雨水、冻融、根系等作用的影响变得疏松,水分入渗能力加强,土壤颗粒之间疏松,容重降低,同时,封育区植被生长良好,植被高度、盖度、地上和地下生物量均提高明显,物种丰富度和多样性也较高[22],土壤结构改善,也利于土壤容重的下降,水分更易下渗,提高土壤持水能力[23-24]。这表明封育不仅影响土壤容重提高植被生物生产力,同时,对生物固碳和土壤持水能力的提高有利。
3.2 土壤贮水量影响因素
土壤贮水量受土壤容重、土壤有机质、生物量等多种因素的影响,而土壤表层更易受到温度等的影响。赵中秋等[25]认为不同土地利用类型土壤的自然含水量、持水性能、供水性能和渗透性能均有显著差异。本研究表明封育区长期禁牧,地面枯落物和腐殖质较多,特别是植物生长初期的5月、6月,以及植物生长末期以后枯落物和腐殖质均较多,对地面覆盖度大,郁蔽环境和大量的枯草堆积减少了蒸散,进而使土壤贮水量增加。未封育区由于自然放牧,受家畜啃食践踏影响,土壤孔隙分布的空间格局发生变化,土壤总孔隙减少,渗透阻力加大[26],影响土壤重力水分的下渗,植被生物量和对地面的覆盖度都较封育低[27],所产生的郁蔽环境较轻,植物的蒸腾和土壤蒸发增强,故封育区土壤贮水量高于未封育区。封育还提高土壤有机质含量,使土壤的团粒结构增加,稳定性团聚体增加,提高了土壤颗粒间的总孔度,利于提高贮水能力。这也与郭泺等[28]关于森林土壤蓄水能力受森林类型枯落物的厚度、孔隙度、有机质含量、温度、土温、容重等多方面因素的影响,不同森林类型条件下土壤蓄水能力差异明显的结果相一致。说明不同利用方式下土壤水分特性与土壤的其他性质如土壤有机质含量、孔隙度、结构团聚体稳定性、容重等有直接或间接的密切关系。土壤的贮水能力在某种程度上还与降水量呈显著相关,一个地区一定时段内降水量增加,贮水量也增加[29],但是所有变量的增加或减少都不是单一因素作用的结果。贮水量受降水量影响,又与植物生物量、有机物质和土壤质地有关[30]。同样,不同时期封育与未封育措施下,土壤贮水量随深度加深其变化规律不尽相同,相互差异各有不同。整体看,高寒草甸的封育措施对土壤上层的影响较深。
3.3 影响土壤持水能力的其他因素
植被截流的因素不可忽略。候琼等[31]指出,随着覆盖度的增加,植被截留量增多,频繁而量小的降水过程增加了植被截留的几率,使累积截流量占降水量的比例增大。本文表明,封育样地长时间禁牧,植被逐渐得到恢复,地上和地下生物量都有提高,地面覆盖度增加,对降水的截流和拦蓄作用明显。土壤持水力还与植被根系有关系[32],因为植物的根系直接着生在土壤里,所以土壤水分含量的多少直接影响到植物根系的发育,反过来,植被根系也会影响土壤的持水能力。本文试验样地处于高寒地区,昼夜温差大,气候干旱,植被类型主要为矮嵩草等具有大量地下根系的高寒草甸,能保护土壤表层减少水力和风力及冻融作用的侵蚀作用,同时,植被根系的存在能增加土壤孔隙度,尤其死亡的根系经过分解,留下的孔隙使土壤变得更松软,从而使表层容重降低。这也与随着土层深度增加,土壤容重增加,饱和持水量、毛管持水量和毛管孔隙度降低的结果相一致。
3.4 结 论
(1) 经11 a封育后高寒草甸0—40 cm各土壤层次容重在1.08~1.27 g/cm3,比未封育的自然放牧区平均降低11%,并达显著性差异(p<0.05),地表0—10 cm层次最为明显。
(2) 与未封育相比,封育11 a后土壤饱和持水量、毛管持水量和田间持水量均有所提高,0—40 cm整层分别增加了6%,9%和11%,封育11 a来分别按1.03,1.37,1.38 mm/a的速率增加。
(3) 封育与未封育条件下土壤贮水量季节动态变化与降水量基本一致,受季节等不同因素影响可分为融冻集聚期、波动消耗期和蓄积期。贮水量在5月底和7月中下旬达到最大值,其中封育和未封育土壤平均贮水量分别为112.58 mm和96.26 mm,表现为封育>未封育。说明封育能促使土壤贮水量增加,利于土壤涵养水分,放牧使土壤贮水量减少。
(4) 土壤贮水量与土壤容重呈极显著负相关,分别与地下生物量、地下生物量呈极显著、显著的正相关,与有机碳密度无相关。
(5) 土壤贮水量在垂直方向上随着土层深度增加呈下降趋势,受降水量影响较大。7月土壤贮水量达到最大值,尤以0—10 cm最为显著,远大于30—40 cm土壤层次。说明不同样地类型土壤贮水量不仅在时间尺度上动态变化,在垂直方向上也存在变化,且在表层和深层封育大于未封育地区。
参考文献:
[1]王文颖,王启基,王刚.高寒草甸土地退化及其恢复重建对土壤碳氮含量的影响[J].生态环境,2006,15(2):362-366.
[2]李媛媛,董世魁,李小艳,等.围栏封育对黄河源区退化高寒草地植被组成及生物量的影响[J].草地学报,2012,20(2):275-279.
[3]刘纪远,徐新良,邵全琴.近30年来青海三江源地区草地退化的时空特征[J].地理学报,2008,63(4):364-376.
[4]邵全琴,樊江文.三江源区生态系统综合监测与评估[M].北京:科学出版社,2012:447-563.
[5]何念鹏,韩兴国,于贵瑞.长期封育对不同类型草地碳贮量及其固持速率的影响[J].生态学报,2011,31(15):4270-4276.
[6]Wu G L, Liu Z H, Zhang L, et al. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China[J]. Plant & Soil, 2010,332(1/2):331-337.
[7]Bilotta G S, Brazier R E, Haygarth P M. The impacts of grazing animals on the quality of soils, vegetation, and surface waters in intensively managed grasslands[J]. Advances in Agronomy, 2007,94:237-280.
[8]范月君,侯向阳,石红宵,等.封育与放牧对三江源区高寒草甸植物和土壤碳储量的影响[J].草原与草坪,2012,32(5):41-46.
[9]徐翠,张林波,杜加强,等.三江源区高寒草甸退化对土壤水源涵养功能的影响[J].生态学报,2013,33(8):2388-2399.
[10]周印东,吴金水,赵世伟,等.子午岭植被演替过程中土壤剖面有机质与持水性能变化[J].西北植物学报,2003,23(6):895-900.
[11]周华坤,赵新全,赵亮,等.高山草甸垂穗披碱草人工草地群落特征及稳定性研究[J].中国草地学报,2007,29(2):13-25.
[12]中国土壤学会农业化学专业委员会.土壤农业化学常规分析方法[M].北京:科学出版社,1984.
[13]刘晓琴,吴启华,李红琴,等.不同封育年限高寒草甸植被/土壤碳密度及净生态系统CO2交换量的比较[J].冰川冻土,2013,35(4):848-856.
[14]李红琴,乔小龙,张镒锂,等.封育对黄河源头玛多高寒草原水源涵养的影响[J].水土保持学报,2015,29(1):195-200.
[15]吴启华,毛绍娟,刘晓琴,等.牧压梯度下高寒杂草类草甸土壤持水能力及影响因素分析[J].冰川冻土,2014,36(3):590-598.
[16]王洁,贾文雄,赵珍,等.祁连山北坡草甸草原地上生物量与土壤理化性质的关系[J].水土保持研究,2017,24(1):36-43.
[17]刘安花,李英年,张法伟,等.高寒矮嵩草草甸植物生长季土壤水分动态变化规律[J].干旱区资源与环境,2008,22(10):125-130.
[18]张社奇,王国栋,时新玲,等.黄土高原油松人工林地土壤水分物理性质研究[J].干旱地区农业研究,2005,23(1):60-64.
[19]Wu G L, Li Z H, Zhang L, et al. Effects of artificial grassland establishment on soil nutrients and carbon properties in a black-soil-type degraded grassland[J]. Plant and Soil, 2010,333(1/2):469-479.
[20]徐翠,张林波,杜加强,等.三江源高寒草甸退化对土壤水源涵养功能的影响[J].生态学报,2013,33(8):2388-2399.
[21]Su X K, Wu Y, Dong S K, et al. Effects of grassland degradation and re-vegetation on carbon and nitrogen storage in the soils of the Headwater Area Nature Reserve on the Qinghai-Tibetan Plateau, China[J]. Journal of Mountain Science, 2015,12(3):582-591.
[22]孙宗玖,安沙舟,马金昌.围栏封育对草原植被及多样性的影响[J].干旱区研究,2007,24(5):669-674.
[23]Eldridge D J, Wang L, Ruiz-Colmenero M. Shrub encroachment alters the spatial patterns of infiltration[J]. Ecohydrology, 2015,8(1):83-93.
[24]李建兴,何炳辉,梅雪梅,等.紫色土区坡耕地不同种植模式对土壤渗透性的影响[J].应用生态学报,2013,24(3):725-731.
[25]赵中秋,蔡运龙,付梅臣,等.典型喀斯特地区土壤退化机理探讨:不同土地利用类型土壤水分性能比较[J].生态环境,2008,17(1):393-395.
[26]Leung A K, Garg A, Coo J L, et al. Effects of the roots of Cynodon dactylon and Schefflera heptaphylla on water infiltration rate and soil hydraulic conductivity[J]. Hydrological Processes, 2015,29(15):3342-3354.
[27]何晴波,赵凌平,白欣,等.封育和放牧对典型草原地上植被的影响[J].水土保持研究,2017,24(4):247-251.
[28]郭泺,夏北成,倪国祥.不同森林类型的土壤持水能力及其环境效应研究[J].中山大学学报,2005,44(4):327-330.
[29]马宗泰.三江源北部天然草地土壤水分动态变化规律[J].安徽农业科学,2009,37(8):3619-3620.
[30]白晓,张兰慧,王一博,等.祁连山区不同土地覆被类型下土壤水分变异特征[J].水土保持研究,2017,24(2):17-25.
[31]候琼,王英舜,杨泽龙,等.基于水分平衡原理的内蒙古典型草原土壤水动态模型研究[J].干旱地区农业研究,2011,29(5):197-203.
[32]Schenk H. Vertical vegetation structure below: Scaling from root to globe[J]. Progress in Botany, 2005,66(2):341-373.
