MaCaM和MaCAMTA3基因在热处理诱导香蕉抗冷性中的作用
2018-04-09王海波苏新国张昭其庞学群
王海波, 李 璐, 苏新国, 张昭其, 庞学群
(1.广东食品药品职业学院,广东广州 510520; 2.华南农业大学生命科学学院,广东广州 510642;3.华南农业大学园艺学院,广东广州 510642)
香蕉果实在贮藏温度低于12 ℃时即发生冷害,难以采用较低的温度来延长贮运期[1-2]。热处理是提高果蔬抗冷性的最有效的采后处理措施之一[3],但热处理诱导的抗冷机制目前尚不明确。钙调素(calmodulin,CaM)是植物中重要的第二信使,CAMTA(calmodulin-binding transcription activator)/SR (signal responsive transcription activator)是一类广泛存在于真核生物中的含有CaM结合位点的转录因子家族。CaM和CAMTA在植物响应逆境中起调节作用,对香蕉CaM和CAMTA 的深入研究,有助于进一步了解香蕉钙信号系统调节的分子机制,有助于揭示热处理提高香蕉果实抗冷性的作用机理。CaM参与了植物对高温、低温胁迫等的响应,并且在植物细胞对胁迫的适应过程中起重要调节作用[4-5]。热、冷、干旱等多种胁迫都能诱导CAMTA基因上调表达[6-7]。拟南芥CAMTA3能同CBF2(C-repeat binding transcription factor)启动子区域CM2序列(CCGCGT)结合,激活CBF2等基因的表达,从而提高拟南芥的抗冷性[8]。笔者所在课题组的前期研究结果表明,采用52 ℃热水处理香蕉果实3 min,能有效减轻香蕉果实的冷害症状[9-10],但有关CaM和CAMTA在热水处理诱导香蕉抗冷性中的作用目前还不明确。分析CaM和CAMTA基因在香蕉热处理和低温贮藏中的表达情况,探讨这2个基因在热处理诱导香蕉抗冷性中的作用,为有效延长香蕉贮运期提供参考依据。
1 材料与方法
1.1 试验材料及处理
香蕉品种为巴西(Musaspp. AAAGroupcv.cavendish),采自广东省广州市番禺香蕉园,及时运回广东省果蔬保鲜重点实验室。挑选七成饱满、大小均匀、无病虫害及机械损伤的单个香蕉, 先后用0.1%漂白粉和0.05%咪鲜胺锰盐可湿性粉剂各浸泡5 min,晾干后备用。
热水及低温贮藏处理:将香蕉果实浸入热水处理机(体积400 L)中,温度52 ℃,时间3 min。热水处理后将香蕉果实放入20 ℃恒温箱中贮藏3 h,然后置于7 ℃下贮藏5 d;对照处理的香蕉在20 ℃恒温箱中贮藏3 h,然后置于7 ℃下贮藏5 d。每个处理100个香蕉,定期取样备用。取样时间点为:20 ℃下0、0.5、1.5、3.0 h;7 ℃下1.0、4.0、8.0、24.0、48.0、120.0 h。
1.2 试验方法
1.2.1脯氨酸含量测定参照Demiral等的试验方法[11],略有改动。取香蕉果皮0.5 g,用5 mL 3%磺基水杨酸提取,沸水浴10 min,冷却。5 000 r/min离心10 min,取上清液待测。测定:取2 mL上清液,加2 mL水、2 mL 3%磺基水杨酸、2 mL冰乙酸和4 mL 2.5%茚三酮,沸水浴显色30~60 min,冷却。加4 mL甲苯萃取,静置后取甲苯相测定520 nm下的吸光度(以甲苯为空白对照)。根据下式计算脯氨酸含量:单位鲜质量样品中脯氨酸含量(μg/g)=2.5C/W,式中:C为样品脯氨酸浓度,μg/mL;W为样品质量,g;2.5表示溶液体积为 2.5 mL。
1.2.2实时定量引物设计根据已经克隆得到的MaCaM和MaCAMTA3基因序列,设计实时定量引物,验证后备用。引物序列为MaCaM(5′-ACCGAAGCTGAGCTACAG-3′、5′-ATCCTTCATCTTGCGAGC-3′)和MaCAMTA3(5′-GCAA ATGGTCACAAAGGC-3′、5′-GAGAAGCATTCCGAACAG-3′)。
1.2.3实时荧光定量PCR(qRT-PCR)采用TOYOBO THUNDERBIRD SYBR®qPCR Mix 进行qRT- PCR扩增。反应体系为20.0 μL,即SYBR-Green PCR Master Mix 10.0 μL,上、下游引物(10 μmol/L)各0.25 μL,稀释的cDNA 2.0 μL,ddH2O 7.5 μL。扩增程序:95 ℃ 3 min;95 ℃ 10 s,59 ℃ 10 s,72 ℃ 20 s,40个循环。为了确认产物的质量和引物的特异性,在溶解曲线中分析产物的Tm(分析溶解温度为65~95 ℃)。所有qRT-PCR结果根据内参基因Actin1进行Ct值校准,再利用2-ΔΔCt计算目的基因的相对表达量,重复3次。
1.3 数据处理
采用Sigmaplot 12.5制作折线图。
2 结果与分析
2.1 热激处理及热激后低温贮藏对脯氨酸含量的影响
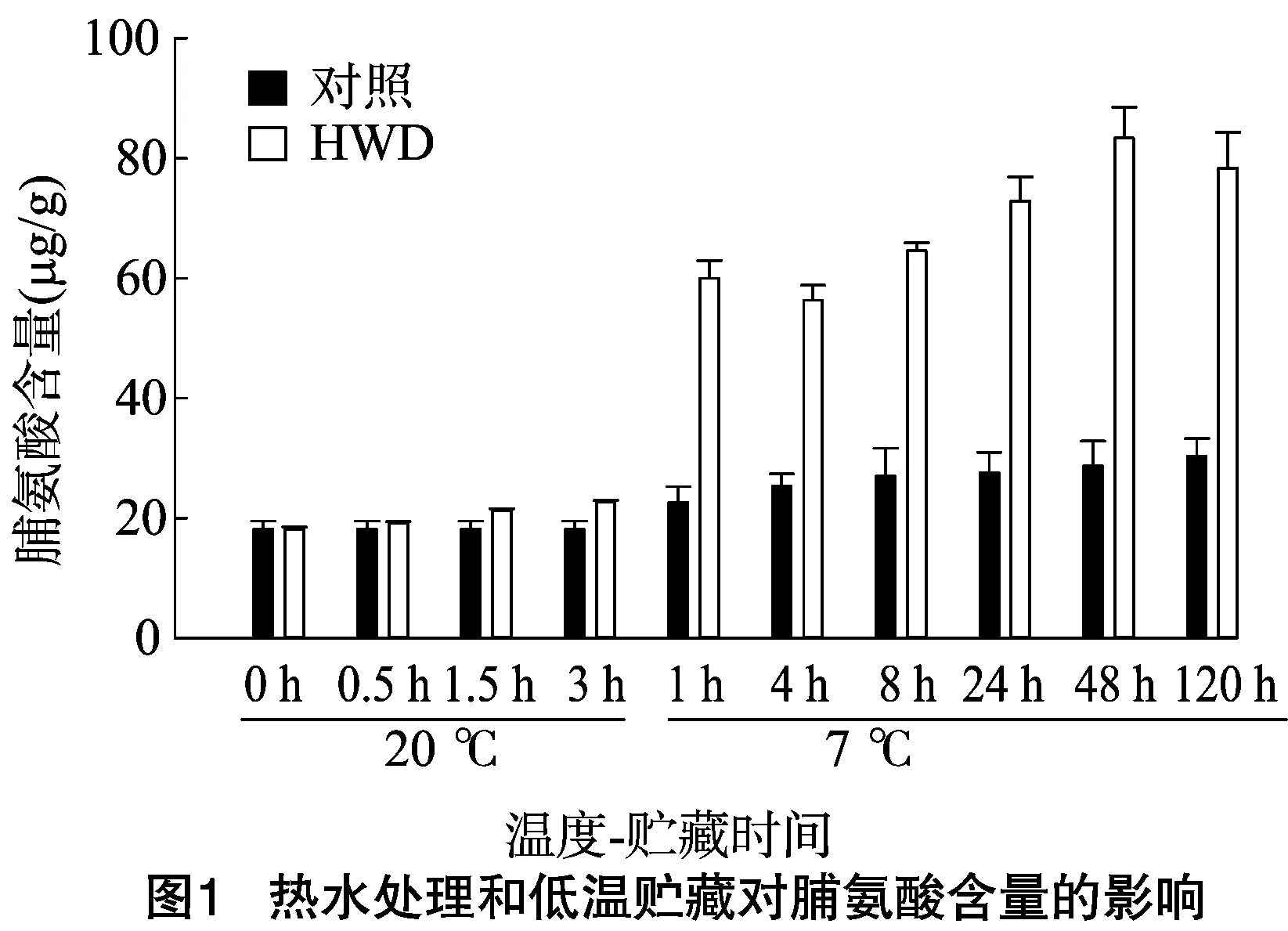
前期研究表明,7 ℃(冷害温度)下,香蕉果实在处理后 5 d 已经出现明显的冷害症状,利用52 ℃热水处理(HWD) 3 min 能有效提高香蕉果实在7 ℃低温贮藏中的抗冷性[9-11]。从图1可知,对照香蕉果实的脯氨酸含量在7 ℃下呈现缓慢上升的趋势。热激处理后在20 ℃贮藏期间,香蕉果实的脯氨酸含量小幅度逐渐增加,且脯氨酸含量略高于对照。当热激后的香蕉果实转入7 ℃下1 h时,脯氨酸含量迅速增加至20 ℃贮藏时的3倍左右,之后呈现出下降上升再下降的趋势,在整个7 ℃低温贮藏期间,热激处理的香蕉果实其脯氨酸含量一直明显高于对照。
2.2 热激处理及热激后低温贮藏对MaCaM表达的影响
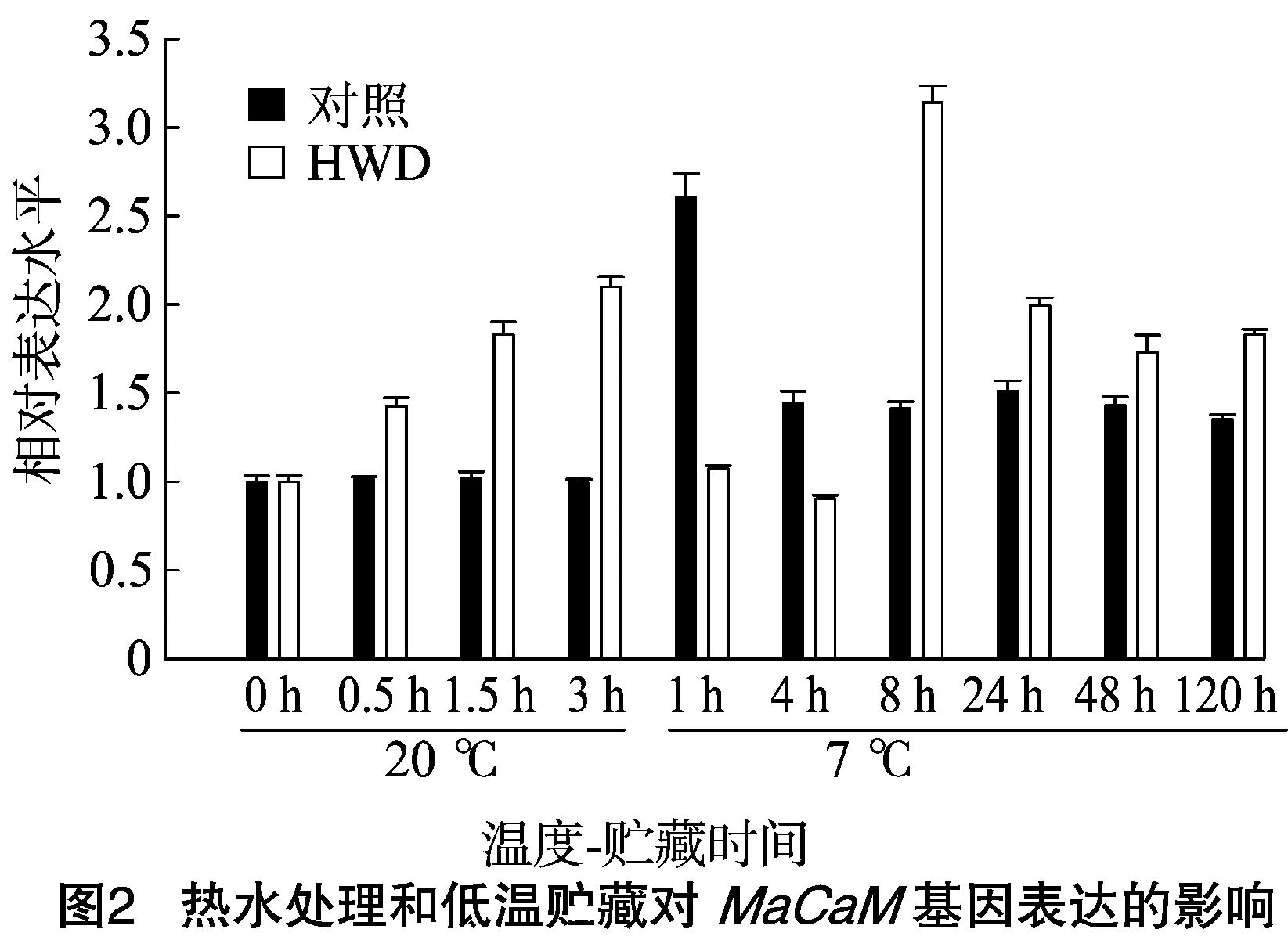
从图2可知,对照处理的MaCaM基因表达在7 ℃下1 h迅速升高至最大值,之后又在7 ℃ 4 h迅速下降,后期持续在同一个水平,但表达量仍高于20 ℃下的表达量。香蕉果实经过HWD处理后,MaCaM基因表达呈现快速升高的趋势,在20 ℃下3 h达到一个高峰,MaCaM基因表达量约为对照的2倍。当热激后的香蕉果实中转入7 ℃低温贮藏1 h时,MaCaM基因表达量迅速下降至0 d的水平,这与对照的表现相反。7 ℃低温4~8 h时,MaCaM基因表达量迅速增强至最高峰,之后逐渐下降。除7 ℃贮藏1、4 h外,热激处理的MaCaM基因表达量均高于对照。
2.3 热激处理及热激后低温贮藏对MaCAMTA3表达的影响
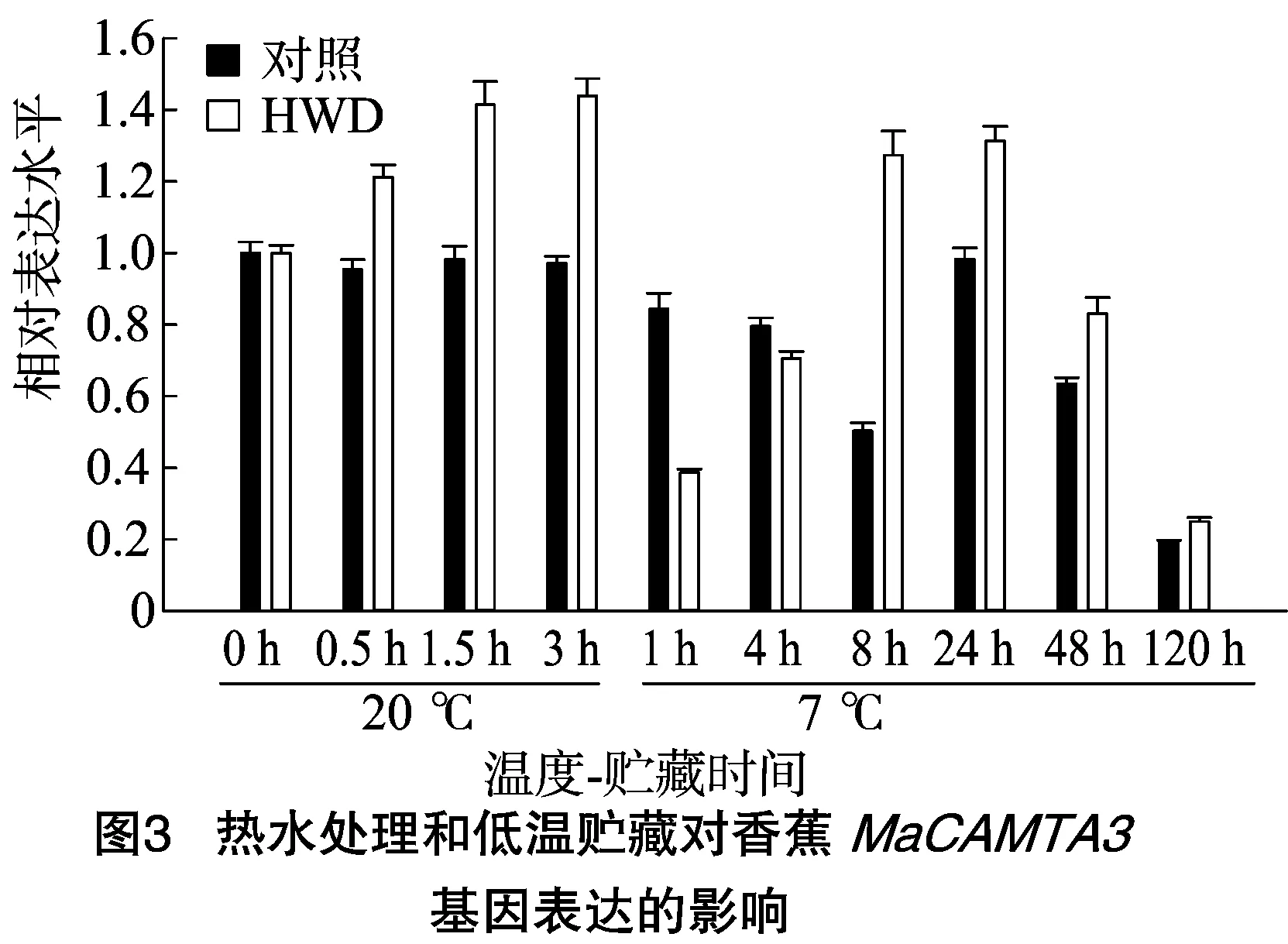
从图3可知,对照处理的MaCAMTA3基因在7 ℃贮藏下1~8 h呈现逐渐下降趋势,7 ℃贮藏8~24 hMaCAMTA3基因表达又增强至20 ℃贮藏时的水平,之后又开始逐渐下降。香蕉经HWD处理后3 h,MaCAMTA3基因表达逐渐升高至最大值,约是对照处理的1.4倍,然而当果实转入7 ℃后1 h,MaCAMTA3基因表达量出现短暂而迅速的下降,且低于对照。之后其表达量逐渐上升,在7 ℃ 8~24 h达到一个高峰,之后又开始逐渐下降。除7 ℃贮藏1、4 h外,HWD处理的MaCAMTA3基因表达均高于对照。
3 结论与讨论
脯氨酸含量的积累与果实抗冷性的提高密切相关[12]。桃果实中较高的脯氨酸含量明显提高了桃果实的抗冷性,而且用外源5 mmol/L脯氨酸处理桃果实,能明显降低桃果实冷害指数,缓解其冷害症状的发生[13]。10 mmol/L甜菜碱处理可提高黄瓜果实中脯氨酸含量,从而减轻黄瓜果实贮藏期间的冷害[14]。外源NO处理能够通过促进香蕉果实脯氨酸积累和增强体系的抗氧化性,进而减轻香蕉冷害[15]。本研究也发现,热激能在处理后3 h内轻微增加香蕉果实脯氨酸的含量,而当果实经热激处理后在7 ℃低温贮藏时,其脯氨酸含量一直明显高于对照处理,说明热激诱导的脯氨酸大量积累与香蕉果实的抗冷性密切相关。
热激处理能在短时间内迅速激活CaM的表达。例如,拟南芥幼苗经过37 ℃热激后,其AtCaM3基因表达水平在 20 min 内迅速增加并达到最大值,在30 min后逐渐下降[16]。37 ℃处理玉米幼苗后,其CaM1-2的表达在10 min后开始迅速增强[4],本研究也发现,香蕉果实经过热激处理后,MaCaM基因表达呈现快速升高的趋势,在处理后3 h达到一个高峰。CaM也参与了植物的低温胁迫反应。例如,茶树在4 ℃低温胁迫下,CsCaM1与CsCaM2基因的表达量从3 h开始逐渐上升,于24 h达到最大值[17]。 4 ℃低温能诱导小麦幼苗TaCaM5的表达量在1 h后迅速升高,3 h达到最高峰,之后又迅速下降至处理前的水平[18]。本研究结果与之类似,7 ℃ 低温诱导香蕉果实的MaCaM基因表达在1 h迅速升高至最大值,之后又在4 h迅速下降,后期维持在一个较低的水平。而经热激处理后在7 ℃低温贮藏的香蕉果实,MaCaM基因表达高峰推迟出现在4 h,除低温贮藏1、4 h外,热激处理的MaCaM基因表达均高于对照。以上结果说明,MaCaM可能参与了热激处理诱导香蕉果实抗冷性的过程。
热处理和低温都能诱导CAMTA基因的表达。42 ℃热处理能诱导拟南芥幼苗的AtCAMTA1、AtCAMTA2、AtCAMTA3、AtCAMTA5、AtCAMTA6这5个基因在15 min开始上调表达,尤其是AtCAMTA1、AtCAMTA5、AtCAMTA6这3个基因在热处理4 h内均维持较高的表达水平。类似地,4℃低温也能诱导拟南芥幼苗AtCAMTA1、AtCAMTA2、AtCAMTA3、AtCAMTA5、AtCAMTA6这5个基因的上调表达,尤其是AtCAMTA1、AtCAMTA3、AtCAMTA5、AtCAMTA6这4个基因在低温处理4 h内均保持较高的表达水平[6]。本研究也发现,热处理能诱导香蕉果实MaCAMTA3基因表达在处理后3 h内迅速增强,7 ℃ 低温贮藏下MaCAMTA3基因表达呈现先下降后上升再下降的趋势,而经过热激处理后在7 ℃低温贮藏的香蕉果实,除低温贮藏1、4 h外,MaCAMTA3基因表达均高于对照。说明热激处理诱导MaCAMTA3基因较高的表达水平,可能参与了热激处理诱导香蕉果实产生的抗冷性。
热处理能诱导香蕉果实在低温贮藏中的脯氨酸含量升高,从而提高果实抗冷性。MaCaM和MaCAMTA3基因可能参与了热激处理诱导香蕉果实产生的抗冷性。
参考文献:
[1]陆旺金,张昭其,季作梁. 热带亚热带果蔬低温贮藏冷害及御冷技术[J]. 植物生理学通讯,1999,35(2):158-163.
[2]张昭其,庞学群. 南方水果贮藏保鲜技术[M]. 南宁:广西科学技术出版社,2008:26.
[3]王海波,张昭其,邓鸿铃,等. 热处理提高采后果蔬抗冷性的机理分析[J]. 广东农业科学,2015,42(15):57-64.
[4]Liu H T,Li B,Shang Z L,et al. Calmodulin is involved in heat shock signal transduction in wheat[J]. Plant Physiology,2003,132(3):1186-1195.
[5]Yang N,Peng C,Cheng D,et al. The over-expression of calmodulin from antarctic notothenioid fish increases cold tolerance in tobacco[J]. Gene,2013,521(1):32-37.
[6]Yang T,Poovaiah B W. A calmodulin-binding/CGCG box DNA-binding protein family involved in multiple signaling pathways in plants[J]. Journal of Biological Chemistry,2002,277(47):45049-45058.
[7]Pandey N,Ranjan A,Pant P,et al. CAMTA1 regulates drought responses inArabidopsisthaliana[J]. BMC Genomics,2013,14(1):216.
[8]Doherty C J,van Buskirk H A,Myers S J,et al. Roles forArabidopsisCAMTA transcription factors in cold-regulated gene expression and freezing tolerance[J]. Plant Cell,2009,21(3):972-984.
[9]王海波,庞学群,黄雪梅,等. 活性氧在热处理诱导香蕉耐冷性中的作用[J]. 中国农业科学,2012,45(5):936-942.
[10]Wang H,Zhang Z,Xu L,et al. The effect of delay between heat treatment and cold storage on alleviation of chilling injury in banana fruit[J]. Journal of the Science of Food and Agriculture,2012,92(13):2624-2629.
[11]Demiral T,Türkan I. Comparative lipid peroxidation,antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance[J]. Environmental and Experimental Botany,2005,53(3):247-257.
[12]刘彤彤,蒋欣梅,于锡宏,等. 间歇降温对黄瓜幼苗耐冷相关指标的影响[J]. 江苏农业科学,2016,44(12):195-196,271.
[13]陈智智. 脯氨酸对减轻桃果实冷害的作用及其机理研究[D]. 南京:南京农业大学,2012.
[14]张海英,王有年,韩涛,等. 外源甜菜碱对黄瓜果实冷藏期间延缓冷害的影响[J]. 中国农业科学,2008,41(8):2407-2412.
[15]Wang Y,Luo Z,Du R,et al. Effect of nitric oxide on antioxidative response and proline metabolism in banana during cold storage[J]. Journal of Agricultural and Food Chemistry,2013,61(37):8880-8887.
[16]Zhang W,Zhou R G,Gao Y J,et al. Molecular and genetic evidence for the key role ofAtCaM3 in Heat-Shock signal transduction inArabidopsis[J]. Plant Physiology,2009,149(4):1773-1784.
[17]黄玉婷,钱文俊,王玉春,等. 茶树钙调素基因CsCaMs的克隆及其低温胁迫下的表达分析[J]. 植物遗传资源学报,2016,17(5):906-913.
[18]刘新颖,王晓杰,薛杰,等. 小麦钙调素新亚型TaCaM5的克隆及表达分析[J]. 作物学报,2010,36(6):953-960.
