黑龙江省马铃薯晚疫病菌的交配型和multi-locus基因型分析
2018-04-04张铉哲韩晓旭郭衍锦李媛媛
张铉哲 韩晓旭 郭衍锦 李 璐 李媛媛
(东北农业大学农学院,黑龙江哈尔滨 150030)
中国是马铃薯生产大国,马铃薯已成为继小麦、玉米之后的第三大作物,种植面积和产量居于世界首位(王立 等,2013)。黑龙江省因得天独厚的冷凉气候和地理条件,成为我国马铃薯主产区之一(关红颖,2011)。然而,近几年由于黑龙江省降雨量大等天气原因,马铃薯晚疫病发生十分严重。
马铃薯晚疫病是由致病疫霉Phytophthora infestans引起的马铃薯毁灭性病害,可造成巨大的经济损失(金光辉 等,2017)。马铃薯晚疫病菌通过不同交配型菌株之间异宗配合进行有性生殖,有性生殖可产生卵孢子,卵孢子可在土壤中越冬,成为翌年的初侵染源。不同交配型同时存在时说明马铃薯晚疫病菌适应性和变异性增强(闵凡祥 等,2010)。Gallegly和Galindo(1958)在墨西哥中部首先发现了A1和A2交配型。张志铭等(1996)首次在国内报道了A2交配型菌株。朱杰华(2004)首次在黑龙江采集分离的菌株中鉴定出A2交配型,其出现频率仅为2.2%。王腾等(2016)对2011~2013年在黑龙江采集的菌株进行交配型测定,A1交配型占菌株总数的51.63%,其余为A2交配型。这些研究表明晚疫病菌由于不同交配型的出现,可以在田间进行有性生殖和无性繁殖,导致晚疫病菌群体结构发生变化。晚疫病菌群体结构变化对晚疫病害的防治提出了新的挑战,因此研究马铃薯晚疫病菌群体遗传结构变化具有重要意义。
线粒体基因组具有结构简单、无组织特异性等优点,采用检测线粒体多态性方法研究晚疫病菌群体遗传结构变化效果更佳(May & Ristino,2004)。对马铃薯晚疫病菌线粒体DNA(mtDNA)多态性进行研究,逐步成为国内外研究热点。Griffith和Shaw(1998)采用改进PCR-RFLP方法将线粒体单倍型分为Ⅰa、Ⅱa、Ⅰb、Ⅱb。Rojas和Kirk(2016)研究表明,2008~2010年在密歇根州采集分离的124株致病疫霉的mtDNA单倍型均为Ⅰa型。Tian等(2016)鉴定了中国西北地区采集分离的马铃薯晚疫病菌mtDNA单倍型,结果检测到Ⅰa、Ⅱa和Ⅱb 3种单倍型,其中Ⅰa为优势基因型。徐生军(2010)利用PCR-RFLP方法,对黑龙江省采集分离的马铃薯晚疫病菌进行mtDNA单倍型检测,共检测到Ⅰa和Ⅱa两种单倍型。可见,4种单倍型在不同国家和地区的分布具有一定差异。
SSR标记具有多态性高、重复性好、数量丰富、发生频率高、批量操作等优点(李建武,2011),近些年成为研究马铃薯晚疫病菌群体遗传多样性的主要方法。对采自黑龙江的致病疫霉进行SSR基因型分析,吴艳清等(2012)鉴定出8种基因型,F-01型所占比例最高。而王晨(2013)的鉴定结果表明,从黑龙江采集的菌株共鉴定出6种SSR基因型,F-01为优势基因型。这些研究表明,黑龙江省马铃薯晚疫病菌的SSR基因型呈现一种动态变化。
Runno-Paurson等(2016)对爱沙尼亚的马铃薯晚疫病菌进行交配型和SSR基因型测定,结果表明该地区出现不同交配型菌株,SSR基因型也趋于多样化。Rekad等(2017)通过SSR标记分析、交配型和瑞毒霉敏感性的测定,对阿尔及利亚西北部地区的致病疫霉进行系统分析,结果表明不同交配型的菌株其对应的优势SSR基因型有差异。Guo等(2010)通过交配型、同工酶基因型、mtDNA单倍型和RG57指纹的分析,发现中国地区的马铃薯晚疫病菌基因型和表现型与其他国家有差异。
我国缺少对马铃薯晚疫病菌multi-locus基因型的研究。本试验首先测定黑龙江省马铃薯晚疫病菌的交配型;其次,利用PCR-RFLP方法和SSR标记测定黑龙江省马铃薯晚疫病菌的mtDNA单倍型和SSR基因型;第三,利用mtDNA单倍型和SSR基因型确定黑龙江省马铃薯晚疫病菌的muti-locus基因型;最终,分析交配型与muti-locus基因型的相互关系。以期为马铃薯晚疫病菌的群体遗传研究和马铃薯晚疫病的防治提供理论依据。
1 材料与方法
1.1 供试菌株及培养基
2015~2017年,在黑龙江省齐齐哈尔市、绥化市、哈尔滨市各马铃薯主产区采集具有马铃薯晚疫病主要症状的单病斑叶片,将马铃薯切成约0.5 cm的薯片,用70%酒精将切后的薯片消毒,将病叶放入灭菌后的选择性培养基中,把消毒处理后的薯片放在病叶上,加入一定的蒸馏水,用封口膜封盖,19 ℃黑暗培养箱中培养4~6 d。
供试培养基为20%番茄汁琼脂培养基和选择性培养基(张铉哲和徐生军,2010)。
1.2 马铃薯晚疫病菌的交配型测定
交配型的测定采用对峙培养法,使用20%番茄汁琼脂培养基。将标准菌株A1(DN-3085日本)、A2(TBC-3韩国)与待测菌株预培养,然后将标准菌株和待测菌株菌饼分别与A1、A2标准菌株对峙培养,菌饼之间距离为3 cm。封盖,每个菌株重复测定3次,19 ℃恒温培养14 d,在显微镜下观察菌落交界处是否产生卵孢子。若待测菌株与A2标准菌株之间产生卵孢子,且与A1标准菌株间不产生卵孢子,说明所测菌株是A1交配型;相反,则是A2交配型。若与2个标准菌株间都产生卵孢子,且自身也可产生卵孢子,则待测菌株类型为自育型。
1.3 DNA提取
将待测菌株接种到20%番茄汁琼脂培养基中,每株菌株接种3皿,19 ℃黑暗培养箱中培养14 d左右,用灭过菌的滤纸吸干菌丝上的水分,装入1.5 mL EP管中,-20 ℃下保存用于DNA的提取。参考Goodwin等(1992)的方法提取基因组DNA。
1.4 马铃薯晚疫病菌mtDNA单倍型测定
1.4.1引物序列与扩增体系线粒体DNA单倍型分析的PCR-RFLP方法参考Griffith和Shaw(1998) 的 方 法。 引 物 序 列:P2:F2:5′- TTCCCTTTGTCCTCTACCCAT-3′,R2:5′-TTACGGCGGTTTAGCACATACA-3′;P4:F4:5′-TGGTCATCCAGAGGTTTATGTT-3′,R4:5′-CCGATACCGATACCAGCACCAA-3′。所用引物由生工生物工程(上海)股份有限公司合成。PCR扩增体系和程序参考Zhang等(2006)的方法。
1.4.2酶切体系目的片段回收采用核酸回收试剂盒〔生工生物工程(上海)股份有限公司〕。P2引物扩增片段使用MspⅠ酶切,P4引物扩增片段使用EcoR Ⅰ酶切,酶切体系为50 μL:胶回收产物10 μL,Buffer 5 μL,Enzyme 1 μL,ddH2O 补足至 50 μL。反应条件为37 ℃水浴3 h。将酶切产物与溴酚蓝混合,采用1.5%琼脂糖凝胶电泳检测,用凝胶成像仪拍照。
1.5 马铃薯晚疫病菌SSR基因型测定
1.5.1SSR引物序列与扩增体系马铃薯晚疫病SSR基因型划分标准参考Knapova和 Gisi(2002) 的 方 法。 引 物 序 列 Pi4B:F:5′-AAAATAAAGCCTTTGGTTA-3′,R:5′-GCAAGCGAGGTTTGTAGATT-3′;Pi4G:F:5′-CGCTGTGTGGATGACAAGTA-3′,R:5′-TCGACCTGACATACGAGCTA-3′。SSR 扩 增 体系和扩增程序参考王晨(2013)的方法。
1.5.2聚丙烯凝胶电泳检测将扩增后的样品与同体积的Loading Buffer混合,95 ℃变性10 min,扩增产物用12%的变性聚丙烯酰胺凝胶电泳,银染染色,用凝胶成像系统拍照(Gene Company Limited GBOX-CHEMI-XRQ)。
2 结果与分析
2.1 马铃薯晚疫病菌分离
2015~2017年在黑龙江省哈尔滨市、绥化市和齐齐哈尔市各马铃薯主产区共采集分离了126株马铃薯晚疫病菌菌株(表1)。

表1 黑龙江省马铃薯晚疫病菌菌株信息
2.2 马铃薯晚疫病菌交配型分析
在黑龙江省采集分离的126株马铃薯晚疫病菌菌株中共发现了3种交配型。A1、A2和自育型菌株分别占所分离菌株总数的88.1%、6.3%和5.6%。从采集年份分析结果显示,2015年采集的22株菌株中,共鉴定出A1和A2 2种交配型,其中,21株菌株为A1交配型(95.5%),仅1株菌株为A2交配型(4.5%)。2016年采集的32株菌株中共鉴定出3种交配型,其中26株A1交配型菌株(81.3%),4株A2交配型(12.5%),2株自育型(6.2%)。2017年共分离鉴定了72株菌株,3株菌株为A2交配型(4.2%),5株菌株为自育型(6.9%),其余64株菌株为A1交配型(88.9%),由此可见,自育型菌株比例逐年增长,各年份之间A1交配型所占比例具有差异。
从采集地区分析结果显示,哈尔滨市采集分离的菌株均为A1型(100%),绥化市采集分离的菌株共鉴定出3种交配型,56株为A1交配型菌株(88.9%),5株为A2交配型菌株(7.9%),2株为自育型菌株(3.2%)。齐齐哈尔市采集分离的菌株共测定出A1、A2和自育型3种,分别为46株(86.8%)、2株(3.8%)和5株(9.4%)。以上结果显示,哈尔滨市马铃薯主产区与其他2个马铃薯主产区相比晚疫病菌交配型相对单一,各地区之间不同交配型所占百分比具有明显差异。
2.3 马铃薯晚疫病菌mtDNA单倍型分析
黑龙江省采集分离的126株马铃薯晚疫病菌菌株共鉴定出Ⅰa和Ⅱa 2种mtDNA单倍型,其中104株菌株为Ⅱa单倍型(82.5%),22株菌株为Ⅰa单倍型(17.5%)。从采集年份分析结果显示,2015年测定的22株菌株中,共测定出19株Ⅱa单倍型菌株(86.4%),3株为Ⅰa单倍型菌株(13.6%)。2016年分离鉴定的32株菌株中,24株为Ⅱa单倍型菌株(75.0%),其余8株为Ⅰa单倍型菌株(25.0%)。2017年分离的72株菌株中,11株为Ⅰa单倍型菌株(15.3%),61株为Ⅱa单倍型菌株(84.7%)。2015年所鉴定菌株中Ⅱa单倍型比例最高。
从采集地区分析结果显示,哈尔滨市采集分离的菌株均为Ⅱa单倍型(100%),绥化市分离的63株菌株中,共鉴定出Ⅰa和Ⅱa 2种单倍型,分别为15株(23.8%)和48株(76.2%)。齐齐哈尔市鉴定的53株菌株中,46株为Ⅱa单倍型(86.8%),7株为Ⅰa单倍型(13.2%)。可见哈尔滨市马铃薯主产区mtDNA单倍型单一,各地区马铃薯晚疫病菌不同mtDNA单倍型所占比例具有明显差异。
2.4 马铃薯晚疫病菌SSR基因型分析
对黑龙江省采集分离的126株马铃薯晚疫病菌菌株进行SSR基因型分析,共鉴定出7种基因型,分 别 为 F-01、F-02、F-03、F-05、F-06、D-02和G-02。其中F-01基因型菌株共鉴定出98株(77.8%),F-05基因型为12株(9.5%),F-03基因型菌株为7株(5.5%),D-02基因型菌株为4株(3.2%),F-06基因型菌株为3株(2.4%),F-02基因型和G-02基因型分别鉴定出1株(0.8%)。
从采集年份来看(图1),2015年分离鉴定的22株菌株中,共鉴定出3种基因型,分别为F-01、F-05和F-03。2016年共鉴定出5种基因型,分别为F01、F02、F03、F05、D02,其中F-01基因型菌株24株(75.0%),占比最高。2017年共鉴定出6种基因型。可见黑龙江省马铃薯晚疫病菌SSR基因型多样性逐年增加,F-01基因型为优势基因型,各年份间F-01基因型所占比例存在差异(图1)。

图1 采集年份与马铃薯晚疫病菌SSR基因型关系
从采集地点来看(图2),哈尔滨市采集分离的菌株只有F-01和D-02 2种基因型,分别占鉴定总菌株数的90.0%和10.0%。绥化市采集分离的菌株SSR基因型最多,有7种,其中F-01基因型47株(74.6%),占比最高。齐齐哈尔市共鉴定出5种SSR基因型(图2)。

图2 采集地区与马铃薯晚疫病菌SSR基因型关系
2.5 马铃薯晚疫病菌multi-locus基因型分析
根据mtDNA单倍型和SSR基因型,2015~2017年黑龙江省采集分离的菌株共划分成9种multilocus基因型(表2),优势基因型均为multi-locus A基因型。其中2015年采集分离的马铃薯晚疫病菌共鉴定出4种multi-locus基因型,2016年共鉴定出7种multi-locus 基因型,其中multi-locus F基因型只在2016年分离的菌株中出现。2017年共鉴定出8种基因型。Multi-locus H基因型和multi-locus I基因型只在2017年分离的菌株中出现。Multi-locus基因型多样性呈逐年增加的趋势。
从表3可以看出,哈尔滨市采集分离的菌株鉴定出3种基因型,绥化市采集分离的菌株共鉴定出9种基因型,其中multi-locus F和multi-locus I基因型仅在绥化市出现。齐齐哈尔市采集分离的菌株中共鉴定出7种基因型。表明绥化市各马铃薯主产区的马铃薯晚疫病菌基因型更为丰富(表3)。
综合上述multi-locus基因型和交配型结果,multi-locus基因型为A基因型的马铃薯晚疫病菌其交配型类型为A1、A2和自育型,基因型B和基因型D的马铃薯晚疫病菌其交配型类型为A1和A2。其余几种multi-locus基因型的马铃薯晚疫病菌交配型都为A1。

表2 黑龙江省分离的马铃薯晚疫病菌采集年份与multi-locus基因型分析
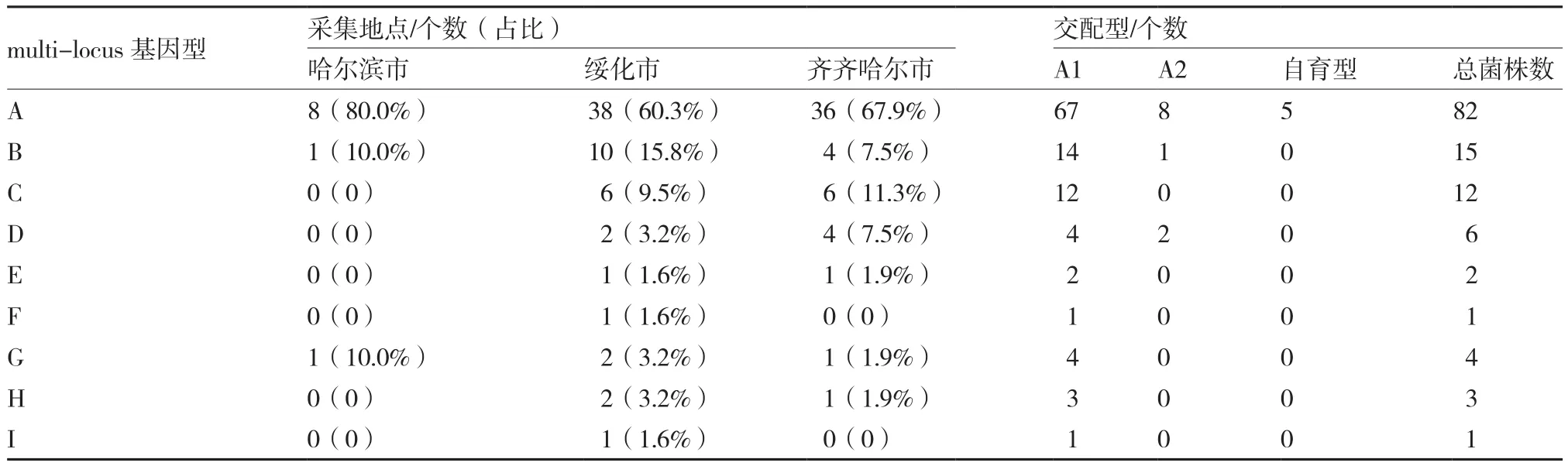
表3 黑龙江省分离的马铃薯晚疫病菌采集地区与multi-locus基因型分析
3 结论与讨论
3.1 马铃薯晚疫病菌交配型
马铃薯晚疫病菌是危害马铃薯、番茄等作物的一种卵菌纲致病真菌,2012~2013年马铃薯晚疫病在全国范围内偏重发生,发生范围广且流行快,对马铃薯造成严重危害(黄冲和刘万才,2016)。
王鹤等(2012)对2009年在黑龙江省采集的菌株进行交配型检测,结果表明鉴定菌株均为A1交配型。郭梅等(2015)研究发现,2011~2012年在黑龙江省哈尔滨、绥化望奎县、齐齐哈尔克山等地发现大量A2交配型马铃薯晚疫病菌菌株,2011年A2交配型占被测菌株的23.08%;2012年占被测菌株的30%。由此可见,黑龙江省马铃薯晚疫病菌交配型逐年复杂,但2014年后有关黑龙江省马铃薯晚疫病菌交配型方面的报道较少。本试验对2015~2017年在黑龙江马铃薯主产区采集分离出的晚疫病菌进行交配型测定,研究发现自育型菌株出现频率逐年增加。从采集地点上发现哈尔滨市只有A1交配型菌株,绥化市和齐齐哈尔市出现3种交配型,说明与其他2个地区相比,哈尔滨市马铃薯晚疫病菌交配型较为单一。这一结论与郭梅等(2015)的研究结果基本一致。同时也说明绥化市和齐齐哈尔市马铃薯主产区有可能发生有性生殖,并导致马铃薯晚疫病菌的遗传多样性更加复杂化。
3.2 马铃薯晚疫病菌mtDNA单倍型和SSR基因型
通过测定马铃薯晚疫病菌基因型,可对晚疫病菌遗传变异进行探究,为防治马铃薯晚疫病提供理论依据。本试验对2015~2017年在黑龙江各马铃薯主产区采集的马铃薯晚疫病菌进行mtDNA单倍型鉴定,鉴定出Ⅰa和Ⅱa两种单倍型。其中,Ⅱa单倍型为优势基因型,与张铉哲等(2015)的研究结果一致,但未发现Ⅰb单倍型菌株。同时,本试验对晚疫病菌进行SSR基因型测定,与李微(2015)对2012~2014年黑龙江省马铃薯晚疫病菌SSR基因型测定结果相比,鉴定出新的基因型F-06基因型。从分离年份上看,2017年黑龙江省晚疫病菌SSR基因型更为丰富,从采集地区方面分析,绥化市各马铃薯产区的马铃薯晚疫病菌SSR基因型更多样化。
3.3 马铃薯晚疫病菌multi-locus基因型
利用马铃薯晚疫病菌的交配型和multi-locus基因型,对马铃薯晚疫病菌的表现型和基因型进行系统综合分析。本试验中,2017年在黑龙江省采集分离的晚疫病菌multi-locus基因型最多,说明黑龙江省马铃薯晚疫病群体结构逐年复杂。从分离地点分析,绥化市采集分离的马铃薯晚疫病菌multi-locus基因型最为丰富。交配型与multi-locus基因型综合分析结果表明,multi-locus A基因型、multi-locus B基因型和multi-locus D基因型与交配型没有相关性,其余multi-locus 基因型与交配型相关。
哈尔滨市各马铃薯主产区马铃薯晚疫病菌multi-locus基因型多样性较低,说明该地区马铃薯晚疫病菌遗传结构较为简单;而绥化市采集的马铃薯晚疫病菌multi-locus基因型最多,表明该地区马铃薯晚疫病菌群体遗传结构更具多样性。总体来说,2015~2017年黑龙江省各马铃薯主产区的晚疫病菌群体结构逐年复杂,给黑龙江省马铃薯晚疫病的防治带来了新的挑战。绥化市可能是黑龙江省马铃薯晚疫病的起源中心,未来该地区应加强对马铃薯晚疫病的监控。
关红颖.2011.黑龙江省马铃薯晚疫病的发生与防治.中国马铃薯,25(3):173-174.
郭梅,Vincent C,闵凡祥,吕军,高云飞,杨帅,王晓丹,Jeanlouis R.2015.黑龙江省发现马铃薯晚疫病菌(Phytophthora infestans)A2交配型.中国马铃薯,(3):171-174.
黄冲,刘万才.2016.近几年我国马铃薯晚疫病流行特点分析与监测建议.植物保护,42(5):142-147.
金光辉,李学湛,王玉成,王腾.2017.年际间干旱对晚疫病菌生理小种复杂性的影响.植物保护,(4):167-173.
李建武.2011.SSR标记技术在马铃薯遗传育种研究中的应用.中国蔬菜,(20):1-8.
李微.2015.黑龙江省马铃薯晚疫病菌群体结构的研究〔硕士论文〕.哈尔滨:东北农业大学.
闵凡祥,王晓丹,胡林双,魏琪,董学志,刘伟婷,郭梅.2010.黑龙江省马铃薯晚疫病菌交配型的研究.中国马铃薯,24(1):47-49.
王晨.2013.马铃薯晚疫病菌的表现型和SSR基因型分析〔硕士论文〕.哈尔滨:东北农业大学.
王鹤,朱杰华,杨志辉,韩彦卿,王宇,徐小虎.2012.2009年黑龙江和吉林省马铃薯晚疫病菌表型结构研究.植物保护,38(1):151-154.
王立,惠娜娜,李建军,马永强,周天旺,李继平,朱小琼,国立耕.2013.甘肃省马铃薯主产区晚疫病菌生理小种组成与分布.中国蔬菜,(11):70-74.
王腾,闵凡祥,郭梅,杨帅,高云飞,金光辉.2016.黑龙江省马铃薯晚疫病菌交配型及对甲霜灵敏感性测定. 植物保护,42(1):180-183.
吴艳清,蒋继志,郑旭,桂春爽,张紫肖,赵磊.2012.中国东北部与河北省致病疫霉SSR基因型的组成与分析.中国农学通报,28(15):170-174.
徐生军.2010.马铃薯晚疫病菌的表现型与基因型的研究〔硕士论文〕.哈尔滨:东北农业大学.
朱杰华.2004.中国马铃薯晚疫病菌群体遗传结构研究〔硕士论文〕.保定:河北农业大学.
张铉哲,徐生军.2010.黑龙江省和吉林省马铃薯晚疫病菌multilocus基因型分析. 中国马铃薯,24(2):97-102.
张铉哲,郝璐,李微,候思宇,赵博,韩晓旭.2015.黑龙江省马铃薯晚疫病菌交配型、瑞毒霉敏感性及mtDNA单倍型分析.吉林农业科学,(5):58-62.
张志铭,李玉琴,田世民.1996.中国发生马铃薯晚疫病菌(Phytophthora infestans)A2交配型.河北农业大学学报,19(4):63-65.
Gallegly M E,Galindo J.1958.Mating types and oospores ofPhytophthora infestansin nature in Mexico.Phytopathology,48:274-277.
Goodwin S B,Spielman L J,Matuszak J M,Bergeron S N,Fry W E.1992.Clonal diversity and genetic differentiation ofPhytophthora infestanspopulations in northern and central Mexico.Phytopathology,82(9):955-961.
Griffith G W,Shaw D S.1998.Polymorphisms inPhytopthora infestans:four mitochondrial haplotypes are detected after PCR.Applied & Environmental Microbiology,64:4007-4014.
Guo L,Zhu X Q,Hu C H.Ristaino J B.2010.Genetic structure ofPhytophthora infestanspopulations in China indicates multiple migration events.Phytopathology,100(10):997-1006.
Knapova G,Gisi U.2002.Phenotypic and genotypic structure ofPhytophthora infestanspopulations on potato and tomato in France and Switzerland.Plant Pathology,51(5):641-653.
May L J,Ristino J B.2004.Identity of the mtDNA haplotype(s)ofPhytophthora infestansin historical specimens from the Irish potato Famine.Mycological Research,108(5):471-479.
Tian Y E,Yin J L,Sun J P,Ma Y F,Wang Q H,Quan J L,Shan W X.2016.Population genetic analysis ofPhytophthora infestansin northwestern China.Plant Pathology,65(1):17-25.
Rekad F Z,Cooke D E,Puglisi I,Randall E,Guenaoui Y,Bouznad Z,Evoli M,Pane A,Lio G M,Cacciola S O.2017.Characterization ofPhytophthora infestanspopulations in northwestern Algeria during 2008-2014.Fungal Biology,5:385-392.
Rojas A,Kirk W W.2016.Phenotypic and genotypic variation in Michigan populations ofPhytophthora infestansfrom 2008 to 2010.Plant Pathology,65(6):1022-1033.
Runno-Paurson E,Kiiker R,Joutsjoki T,Hannukkala A.2016.High genotypic diversity found among population ofPhytophthora infestanscollected in Estonia.Fungal Biology,120:385-392.
Zhang X Z,Kim H Y,Kim B S.2006.Analysis of genetic diversity ofPhytophthora infestansin Korea by using molecular markers.Journal of Microbiology & Biotechnology,16(3):423-430.
