基于PCR-RFLP技术分析H-FABP基因在5个猪群体中的多态性
2018-02-28吴华莉涂尾龙曹建国都启晶常谈永松
吴华莉,涂尾龙,曹建国,都启晶,常谈永松*
(1.上海市农业科学院畜牧兽医研究所,上海 201106;2.上海市调控生物学重点实验室,上海 200241;3.上海种猪工程技术研究中心,上海 201302;4.青岛农业大学食品科学与工程学院,山东 青岛 266109;5.云南农业大学动物科学技术学院,云南 昆明 650201;6.淮安洪泰良种猪繁育有限公司,江苏 淮安 223472)
【研究意义】猪肉的品质和口感与猪肉的嫩度、风味和多汁性相关,而存在于肌纤维束周围结缔组织中的肌内脂肪(IMF)影响着猪肉嫩度。一般认为IMF 达到2 %较为适宜。猪肉IMF的沉积与遗传变异有关。【前人研究进展】脂肪酸结合蛋白家族(FABPs)能够结合细胞内的脂肪酸,是运输细胞内脂肪酸最重要的载体,也有研究表明FABPs有可能参与细胞内脂肪酸双向转运过程[1]。心脏型脂肪酸结合蛋白(heart fatty acid binding protein,H-FABP)是FABPs中的一员。H-FABP基因不仅在各个组织都有表达,尤其在心肌和骨骼肌细胞中呈现高表达[2]。Gerbens等发现H-FABP基因是调控猪肉IMF含量的重要候选基因[3]。国内学者发现H-FABP基因与IMF具有极强的相关性[4-5]。【本研究切入点】本研究目的在于检测H-FABP基因在上海地区5个猪群体中的多态性。【拟解决的关键问题】以期为上海地区这5个猪群肉质方面的育种工作提供分子标记。
1 材料与方法
1.1 供试样品
采集组织样品包括:杜洛克(100头)、长白猪(80头)、大白猪(150头)来自上海祥欣畜禽有限公司,申农猪(80头)来自上海富民农场,梅山猪(140头)来自于上海状元猪场,采集部位为耳组织块。使用试验试剂Taq酶购于北京康为世纪生物科技有限公司,HaeШ酶购于宝生物工程有限公司。
1.2 试验方法
1.2.1 DNA提取 采用AXYGEN基因组试剂盒提取组织DNA,测定DNA浓度,稀释DNA,使每管浓度达到30 ng/mL,-20 ℃保存备用。
1.2.2 引物及PCR扩增 上游引物:5’-ATTGCTTCGGTGTGTTTGAG-3’;下游引物:5’-TCAGG AATGGGAGTTATTGG-3’[6]。PCR扩增条件为: 94 ℃预变性4 min; 94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸30 s,共36个循环;最后72 ℃延伸10 min。PCR产物由1 %琼脂糖凝胶电泳检测,要求目的条带清晰、单一无杂带,符合条件的PCR产物可以进行酶切试验,采用1.5 %琼脂糖凝胶电泳检测酶切结果。
1.2.3 基因型分析 猪H-FABP基因PCR扩增片段为816 bp,第2内含子区域存在3处HaeШ酶切位点,其中一个酶切位点存在多态性,具体的基因型判定标准见表1。根据琼脂糖检测酶切结果,不同基因型分别选取3个样品进行测序。基因型判定时需结合琼脂糖检测结果和测序结果进行分析。

表1 H-FABP基因酶切后基因型判定标准
2 结果与分析
2.1 PCR-RFLP检测
用琼脂糖凝胶电泳检测酶切结果,目的条带大小与预期一致。图1显示,DD型为3条带,dd型为4条带,Dd型杂合子为5条带。根据电泳检测结果统计每个个体的基因型。
2.2 H-FABP基因在5个猪群体中的基因型频率和等位基因频率
采用PCR-RFLP技术,在5个猪群体中进行H-FABP基因的多态性检测。具体的基因型频率和等位基因频率见表2。
由表2可见,H-FABP基因各基因型在5个猪群体中的分布情况不同:在杜洛克群体中以dd型为主要基因型,有较少的杂合子Dd的基因型和DD基因型,占优势的是d等位基因;在大白猪群体中占优势的是Dd基因型,D和d等位基因频率均匀分布;在申农猪群体中检测出3种基因型,主要为dd型,d等位基因频率高于D等位基因频率;在长白猪群体中只检测出dd基因型;在梅山猪群体中只检测出DD基因型。本研究结果显示H-FABP基因在杜洛克、大白、申农猪群体中检测出3种基因型,但在长白和梅山猪群体中出现偏态现象。
3 讨 论
H-FABP基因不仅参与骨骼肌胞内脂肪酸的转运,并且在脂类代谢方面发挥重要作用[7]。H-FABP基因突变位点已被证实和杜洛克*梅山猪、杜洛克、杜洛克*长白猪的脂肪性状有关[8]。有研究表明HaeШ酶切位点产生dd型比DD型的H-FABP基因mRNA表达量低,比DD型阉猪的背最长肌IMF高0.24,达到显著水平(P<0.05),比DD型母猪的背最长肌IMF高0.02,影响不显著[2]。本研究在梅山猪群体内只检测出DD型,这与已报道我国地方猪种在此位点缺乏多态性的研究结果一致[9-10]。本研究在长白猪群体内只检测出dd型,这与林万华等检测H-FABP基因在长白群体中检测出3个基因型的结果不同,造成差异的原因①可能与本次样品都为长白母猪有关,②采样猪种来源不同,但以上推论需要进一步验证。
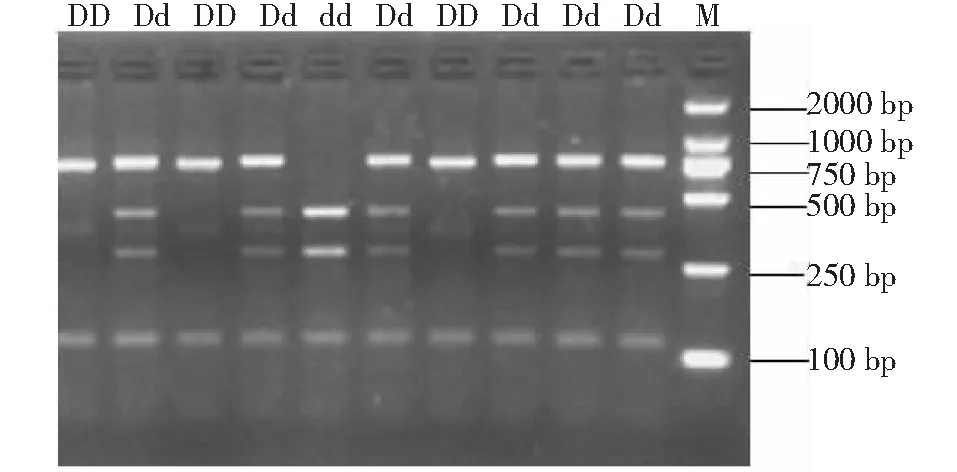
图1 H-FABP基因酶切检测Fig.1 Results of H-FABP gene digested by endonuclease
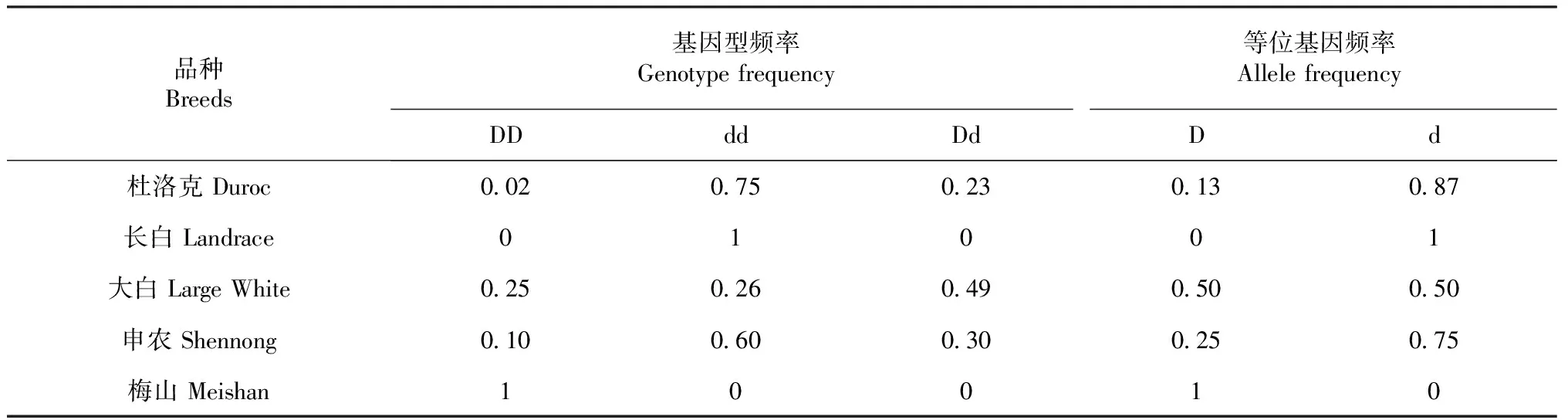
表2 H-FABP基因在5个猪群体中的基因型频率和等位基因频率
此外,研究者发现H-FABP基因5'上游区域存在HinfⅠ酶切位点、第2内含子区域存在MspⅠ和HaeⅢ酶切位点,这些位点与不同猪种IMF相关性并不完全一致[11-12]。研究证实国外猪种3个酶切位点产生“aa-dd-HH”型与猪高肌内脂肪含量相关。检测110 kg体重杜洛克时,发现dd型的IMF含量较高,而DD型猪背膘较低。H-FABP基因多态性影响IMF含量变化原因可能在于能更有效的增加脂肪酸转运或者调节细胞内脂肪酸转运的不同[3]。Pang等也证实dd型脂肪细胞的沉积能力高于DD型[13]。 Chao等研究证实DD型个体瘦肉率高于dd型[14]。本研究检测H-FABP基因在dd型在杜洛克、长白和申农猪群体内频率均大于0.5以上,在育种过程中要关注这3个猪群体DD型个体比例,可改善猪肉合适IMF含量。
有学者研究H-FABP基因的mRNA 表达水平与蛋白质浓度时,发现两者之间相关性较低,说明该基因的表达调节不受转录调控,而受翻译调控。猪体内IMF特性变化与脂肪沉积的生长周期有关,在猪生长过程中观察IMF含量变化,不适当的采集点监测H-FABP基因mRNA 量和蛋白质水平并不能表明IMF的遗传变异情况,通常选择脂肪沉积后180 d日龄屠宰测定[2]。此外,背最长肌的纤维类型有可能也影响H-FABP基因检测结果[15]。
动物育种工作者们关于H-FABP基因mRNA、蛋白质以及不同酶切位点的多态性分析,目的都是为提高或者降低猪肉IMF,改善猪肉品质方面的育种提供辅助遗传标记。本研究的结果可为5个猪群体IMF改良提供基础数据。尤其在实际育种过程发现本实验室选育“长白*大白*二花脸”杂交品系申农猪的脂肪性状不理想,这可能与申农猪群体中H-FABP基因dd型较多有关,接下来可结合申农猪IMF测定结果进行相关分析,期望为该品系肉质改良提供有价值的参考。
4 小 结
本研究对H-FABP基因第2个内含子区域HaeШ酶切位点进行多态分析,确定5个猪群体中该位点的优势基因型,后续可结合IMF性状进一步分析,从而进行标记辅助育种工作。
[1]Zimmerman A W, Veerkamp J H. Members of the fatty acid-binding protein family inhibit cell-free protein synthesis[J]. FEBS Lett, 1998, 437(3): 183-186.
[2]Gerbens F, Verburg F J, Van Moerkerk H T, et al. Associations of heart and adipocyte fatty acid-binding protein gene expression with intramuscular fat content in pigs[J]. J Anim Sci, 2001, 79(2): 347-354.
[3]Gerbens F, van Erp A J, Harders F L, et al. Effect of genetic variants of the heart fatty acid binding protein gene on intramuscular fat and performance traits in pigs[J]. J Anim Sci, 1999, 77(4): 846-852.
[4]Chen J N, Jiang Y Z, Cen W M, et al. Distribution ofH-FABPandACSL4 gene polymorphisms and their associations with intramuscular fat content and backfat thickness in different pig populations[J]. Genet Mol Res, 2014, 13(3): 6759-6772.
[5]Nechtelberger D, Pires V, Söolknet J, et al. Intramuscular fat content and genetic variants at fatty acid-binding protein loci in Austrian pigs[J]. J Anim Sci, 2001, 79(11): 2798-2804.
[6]林万华,任 军,丁能水,等.H-FABP基因型对二花脸猪相关性状影响的初步分析[J].畜牧兽医学报,2003,34(4):318-324.
[7]OVilo C, Oliver A, Noguera J L, et al. Test for positional candidate genes for body composition on pig chromosome6[J]. Genet Sel Evol, 2002, 34(4): 465-479.
[8]Gerbens F, de Koning D J, Harders F L, et al. The effect of adipocyte and heart fatty acid binding protein genes on intramuscular fat and backfat content in Meishan crossbred pigs[J]. J Anim Sci, 2000,78(3): 552-559.
[9]林万华,黄路生,任 军,等. 中外十个猪种H-FABP基因遗传变异的研究[J]. 遗传学报,2002,29(1):12-15.
[10]许卫华,康丽珠,林秀娇,等. 槐猪H-FABP基因多态性及其与肌内脂肪含量相关性研究[J]. 福建师范大学学报(自然科学版),2014, 30(2):76-80.
[11]彭先文,梅书棋,李良华,等.湖北白猪及其杂交后代H-FABP基因型与相关性状关系研究[J].中国农业科学,2008,41(2):570-575.
[12]李志娟,高士争,潘洪彬,等.不同H-FABP基因型滇南小耳猪脂类合成代谢相关基因表达水平研究[J]. 中国细胞生物学学报,2013,35(5):661-667.
[13]庞卫军,白 亮,杨公社. 西部地区主要猪种H-FABP基因多态性、IMF含量及不同基因型脂肪细胞脂滴量的关系[J]. 遗传学报,2006,33(6):515-524.
[14]Chao Z, Wang F, Deng C Y, et al. Distribution and linkage disequilibrium analysis of polymorphisms ofMC4R,LEP,H-FABPgenes in the different populations of pigs, associated with economic traits in DIV2 line[J]. Mol Biol Rep, 2012, 39(5): 6329-6335.
[15]Brocks L, Hulsegge B, Merkus G. Histochemical characteristics in relation to meat quality properties in the longissimus lumborum of fast and lean growing lines of large white pigs[J]. Meat Sci, 1998, 50(4): 411-420.
