盐胁迫对向日葵木质部离子转运的影响
2017-11-30刘杰耿淑娟
刘杰+耿淑娟


摘要:以向日葵品种白葵杂6号为试验材料,中性盐NaCl、Na2SO4按摩尔比9 ∶ 1混合模拟盐胁迫,通过测定其木质部液的成分来探讨盐胁迫对向日葵木质部液中离子转运的影响。结果表明,向日葵的子叶节区对离子的向上运输具有截流作用,尤其是Na+;同时,盐胁迫下,向日葵保持了较高的K+吸收率及贡献率,从而使向日葵体内保持高K低Na状态,这可能是向日葵抗盐性高于其他作物的主要原因。
关键词:向日葵;木质部;盐胁迫;离子转运
中图分类号: S565.501 文献标志码: A 文章编号:1002-1302(2017)20-0085-03
土壤盐渍化已成为全球土壤面临的严重环境问题,而且是制约世界农业经济发展的重要因素[1]。我国土地盐渍化大约占全球盐渍土面积的1/10,从滨海到内陆,从低地到高原都分布着不同类型的盐渍化土地[2]。植物对盐逆境适应能力的研究已成为研究者越来越关注的焦点,如何提高植物的抗盐能力也成为研究的重中之重。
盐胁迫对植物的伤害作用有2个方面,一方面是盐的原初作用,即盐离子的直接毒害作用;另一方面是盐的次生作用,即由盐离子大量积累引起的渗透胁迫和营养匮乏,这2个方面均与植物对盐胁迫离子的吸收和分配有直接或间接的关系[3]。盐胁迫下,植物为维持正常的生理代谢,组织或器官会对离子进行选择性吸收及运输来调控离子的分布,以降低胁迫对自身的伤害[4]。
向日葵是世界上重要的经济作物之一,而且对盐胁迫有一定的耐受力。目前学者对向日葵体内离子情况研究集中于根、叶及子叶整体情况的分析[1,5];而对于盐胁迫下,向日葵木质部汁液中离子变化分析未见报道。本研究从盐胁迫下向日葵子叶节上下木质部汁液中离子变化角度分析,来探讨盐胁迫下,子叶节对向日葵体内离子从地下部分转移到地上部分过程中的作用。
1 材料与方法
1.1 供试材料
向日葵(Helianthus annuus)种子为吉林省向日葵研究所提供的白葵杂6号,以自选不育系H-01A为母本,RHA-K为父本杂交而得,是吉林省向日葵的主要耕种品种。
选择饱满一致的向日葵种子植于直径17 cm的盛满洗净细沙的塑料盆中。等距播种4穴,每穴2粒种子。出苗整齐后,每穴留苗1株,每盆留苗4株。人工遮雨。
1.2 盐碱胁迫条件设计
将2种中性盐NaCl、Na2SO4按摩尔比9 ∶ 1混合,作为盐胁迫(salt stress)组。根据向日葵对中性盐的耐受程度及木质部液的流出程度,设30、60、90 mmol/L 3个处理浓度,分别记为S30、S60、S90。处理液用1倍的Hoagland营养液配制,对照(CK)为1倍的Hoagland营养液。
1.3 胁迫处理
苗龄5周后进行胁迫处理。将长势均匀的向日葵随机分为4组,分别为CK、S30、S60、S90,每个处理6盆。于每天 17:00—18:00用500 mL处理液分2次透灌幼苗,对照组用营养液透灌。处理时间为2 d。
1.4 木质部汁液的收集
将每个处理的6盆幼苗,随机再分成2组。一组用以收集子叶节上的木质部液,另一组用以收集子叶节下的木质部液。收集时用刀片分别在子叶节上下大约2 cm处切割茎,并用硅胶管固定在茎上[6],最后用保鲜膜将硅胶管开口处封上,防止灰尘落入,同时减少蒸发量。收集时间为12 h,收集到的伤流液即为木质部汁液。
1.5 离子测定
K+和Na+含量测定采用火焰分光光度法[7];Cl-、NO3-、H2PO4、SO42-含量测定采用离子色谱法[8];Ca2+、Mg2+含量采用原子吸收分光光度计测定[8]。
1.6 数据统计分析
所有数据均以3次重复的平均值及其标准误差来表示,数据处理及方差计算均用统计学软件SPSS 14.0进行,检验水平均为0.05。其中多重比较采用LSD法。
2 结果与分析
2.1 盐胁迫对向日葵木质部液中离子含量的影响
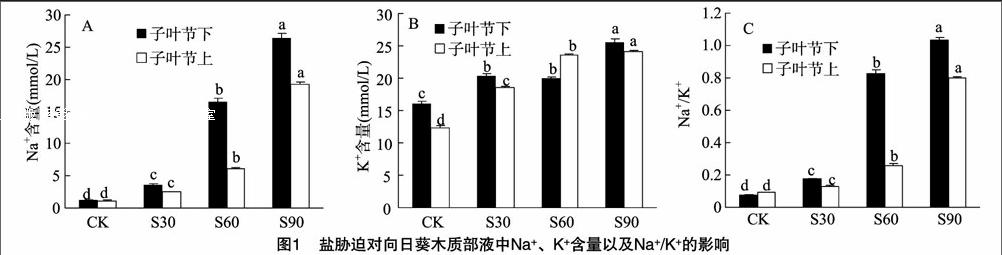
2.1.1 盐胁迫对向日葵木质部液中Na+、K+含量的影响 子叶节上下木质部液中Na+含量均随着胁迫强度增加而显著升高,且子叶节上木质部液中Na+含量低于子叶节下含量,说明Na+在通过木质部向上运输时,浓度逐渐降低(图1-A)。盐胁迫下,子叶节上下木质部液中K+含量随盐度增加而升高,但子叶节上下含量基本相当(图1-B)。盐胁迫下,2个部位木质部液中钠钾比都随胁迫强度增加而有所上升(图1-C)。总体看来,木质部K+含量高于Na+含量。
2.1.2 盐胁迫对向日葵木质部液中阳离子含量的影响 盐胁迫对于木质部液中Mg2+、Ca2+含量影响呈现出低浓度盐胁迫降低其含量、高浓度盐胁迫使其含量增加的趋势;并且在高浓度盐胁迫下,子叶节上木质部液中Mg2+、Ca2+含量远远高于子叶节下木质部液的含量(图2)。
2.1.3 盐胁迫对向日葵木质部液中阴离子含量的影响 由图3可见,子叶痕下木质部液中,Cl-含量随盐胁迫强度的增强逐渐升高,且在盐胁迫强度为30、60 mmol/L时子叶节下木质部液中Cl-含量明显高于子叶节上含量。2个部位相比较,子叶节上的木质部液中Cl-含量較少,子叶节区对Cl-的向上运输具有一定限制作用。木质部液(子叶节下和上)中,NO3-和H2PO4-含量都明显低于对照组,在木质部中向上运输的过程中都有所减少。盐胁迫都使子叶节下木质部液中SO42-含量明显上升,但是子叶节下木质部液中SO42-含量无太大变化。
2.2 盐胁迫对向日葵木质部液电荷贡献率的影响
正常生长的向日葵子叶节下木质部液中,K+电荷贡献率为77.25%,Na+为5.97%,盐胁迫虽然导致子叶节下木质部液中K+贡献比例降低,Na+贡献比例升高,但最终二者贡献比相平;而子叶节上木质部液中K+贡献率始终高于Na+贡献率。endprint
盐胁迫下,Mg2+和Ca2+贡献率比对照有一定程度的下降,但子叶节上木质部液贡献率略大于子叶节下贡献率。盐胁迫下Cl-电荷贡献率快速增加,平均贡献率分别为59.58%(子叶节下)、51.25%(子叶节上)。NO3-所占比例却骤降,子叶节下从对照的80.29%降到盐胁迫下21.67%;子叶节上从81.36%下降到31.52%。SO42-提供负电荷比例随着胁迫强度增加而有所降低,但整体高于对照。H2PO4-贡献率随着胁迫强度的增加而明显降低(表2)。
3 讨论与结论
Na、K、Ca作为植物生长所必需的大量矿质元素,在植物生长发育过程中发挥着重要的生理功能[3]。但是,这些离子只有处于相对平衡状态时才能发挥其正常的生理作用,而离子的稳态平衡一旦破坏将对植物生长及其生理功能产生不利影响[9]。
一般情况下,植物对Na+、K+吸收有拮抗作用。植物为能在盐胁迫环境下生长,会大量吸收土壤中Na+以维持细胞内渗透压;结果会导致K+吸收降低。但在本试验中,木质部液中Na+、K+含量均随着盐胁迫强度的增加而升高,并且由图1可以看出,子叶节上Na+浓度明显低于叶节下Na+浓度,而K+浓度在二者中差异不大,这就说明子叶节对Na+有一定的截留作用,这就使得向日葵地上部分有较高的K+,从而使向日葵体内保持高K低Na状态,从而表现出相对高于其他作物的抗盐性,这与笔者前期试验结果[1,5]相吻合。
盐胁迫下,Ca2+在离子的选择性吸收和保持细胞膜的完整性方面发挥着重要作用[10]。Ca2+在子叶节上木质部液中显著增加,这可能是盐胁迫诱导Ca2+从周围钙库中释放出来,参与茎或叶片生理活动,猜测Ca2+的增加主要起到信号功能传递以及渗透调节的作用。
Mg是植物生长必需的矿质营养元素,杨立飞等认为,盐胁迫下叶片中Mg2+含量升高,有利于增强光合作用,从而提高植株的耐盐性[11]。本研究中,子叶节上木质部液Mg2+含量随盐胁迫的增加呈上升趋势,这表明向日葵根系在盐胁迫会加快Mg2+的吸收并向上运输,以保证叶片的正常光合作用,抵抗外界胁迫的伤害。
盐胁迫导致木质部液中各离子含量及电荷贡献率发生变化,这可能会导致正负电荷失衡,进而影响到木质部液的pH值。研究发现,活体植物乃至离体的叶片,其木质部液的pH值和K+的变化动态(可能也有Na+的变化)都呈显著相关,木质部液流pH值受K+(Na+)离子活性变化的影响[12]。以前的研究认为pH值的改变是一种信号,Wilkinson等用番茄的野生型和突变体研究时发现,pH值升高并非信号,而是逆境下的一种伤害表现[13]。目前,研究者还没有弄清楚木质部液中pH值升高的机制,可能是单一途径调节,也可能是多渠道实现。
综上所述,盐胁迫下,子叶在植物木质部离子准运过程中起了一定作用,尤其是对毒害离子(Na+、Cl-)的向上运输有一定的截留作用,从而保证了地上部的正常生长。
参考文献:
[1]刘 杰,李云玲,孙 虎,等. 盐碱胁迫对向日葵子叶渗透调节及离子平衡的影响[J]. 安徽农业大学学报,2015,42(1):86-91.
[2]张金林,李慧茹,郭姝媛,等. 高等植物适应盐逆境研究进展[J]. 草业科学,2015,24(12):220-236.
[3]李玉梅,郭修武,姜云天. 碱性盐胁迫对牛叠肚幼苗离子积累和运输的影响[J]. 贵州农业科学,2016,44(2):61-66.
[4]陈惠哲,Ladatko N,朱德峰,等. 盐胁迫下水稻苗期Na+和K+吸收与分配规律的初步研究[J]. 植物生态学报,2007,31(5):937-945.
[5]Liu J,Shi D C. Photosynthesis,chlorophyll fluorescence,inorganic ion and organic acid accumulations of sunflower in responses to salt and salt-alkaline mixed stress[J]. Photosynthetica,2010,48(1):127-134.
[6]邹春琴,张福锁. 叶片质外体pH降低是铵态氮改善植物铁营养的重要机制[J]. 科学通报,2003,48(16):1791-1795.
[7]刘 杰,张美丽,张 义,等. 人工模拟盐、碱环境对向日葵种子萌发及幼苗生长的影响[J]. 作物学报,2008,34(10):1818-1825.
[8]刘 杰. 向日葵對碱胁迫和盐胁迫适应机制比较[D]. 长春:东北师范大学,2011.
[9]韩志平,郭世荣,郑瑞娜,等. 盐胁迫对小型西瓜幼苗体内离子分布的影响[J]. 植物营养与肥料学报,2013,19(4):908-917.
[10]夏 阳,梁慧敏,束怀瑞,等. NaCl胁迫下苹果幼树叶片膜透性、脯氨酸及矿质营养水平的变化[J]. 果树学报,2005,22(1):1-5.
[11]杨立飞,朱月林,胡春梅,等. NaCl胁迫对营养液栽培嫁接黄瓜生物量及离子分布的影响[J]. 西北植物学报,2006,26(12):2500-2505.
[12]Felle H H,Herrmann A,Hückelhoven R,et al. Root-to-shoot signalling:apoplastic alkalinization,a general stress response and defence factor in barley (Hordeum vulgare)[J]. Protoplasma,2005,227(1):17-24.
[13]Wilkinson,S,Corlett J A,et al. Effects of xylem pH on transpiration from wild-type and flacca tomato leaves. A vital role for abscisic acid in preventing excessive water loss even from well-watered plants[J]. Plant Physiology,1998,117(2):703-709.endprint
