广东省大沙河水库轮虫群落结构的时空异质性分析
2017-11-10徐少林李慧明顾杨亮韩博平
徐少林, 李慧明, 顾杨亮, 韩博平
暨南大学生态学系,广州 510632
广东省大沙河水库轮虫群落结构的时空异质性分析
徐少林, 李慧明, 顾杨亮, 韩博平*
暨南大学生态学系,广州 510632
于2010年1月至12月对广东省大沙河水库中的河流区、过渡区和湖泊区进行逐月采样, 采集浮游动物并测定水体理化指标, 以探讨热带大型富营养化水库轮虫群落结构的时空异质性。在三个采样点, 形态学分类共鉴定出 25种轮虫,其中19种来自游泳亚目的6个科, 8个属, 4种来自簇轮亚目的1个科, 2个属, 剩余2种来自胶鞘亚目的2个科, 2个属。数据分析表明: 1)大沙河水库轮虫群落结构变化具有明显的季节特征, 可分为枯水一期(1—4月), 丰水期(5—9月)和枯水二期(10—12月), Indval分析显示不同季节的指示种不同且均不少于三种; 2)轮虫群落结构和多样性(Shannon-Hill number)不具有明显的空间异质性, 但轮虫多样性和物种数具有显著的季节差异 (p= 0.004,p= 0.007) ; 3)环境变量对轮虫群落结构的解释率为41.269%, 水体理化变量解释率31.935%, 其中温度解释率16.512%, 水库水文变量对轮虫群落结构变化解释率9.334%,水体理化变量对轮虫群落结构有重要的影响。
轮虫; 群落结构; 时间异质性; 空间异质性; 水体理化变量
1 前言
轮虫是浮游动物的重要类群, 具有世代周期短、种群丰度高等特点, 在自然水体中能够维持较高种群数量。轮虫主要以水体中的颗粒有机物为食物来源(包括藻类、细菌和有机碎屑等), 是水体食物链“上行效应”和“下行效应”中的关键环节, 其种类组成能够较好地反映水体营养水平、理化特征及生境多样性[1]。因此, 了解轮虫的生物群落的组成结构及其时空异质性对水库的水质评价与管理具有重要意义。
在浮游动物食性鱼类较多的水体中, 由于鱼类对浮游动物的捕食通常具有个体大小上的选择性,大型枝角类和桡足类受较强的捕食压力, 而个体较小的轮虫所受捕食压力相对较小, 其丰度和多样性通常较高[2–4]。大型水库具有广阔的水域, 为复杂生境形成与存在提供了条件, 也为不同生态位的轮虫种类提供了共存空间条件[5]。在水体富化营养化过程中, 浮游植物群落多样性下降, 少数浮游植物优势种起主导作用, 导致轮虫的食物质量下降, 进而影响其群落结构[6–8]。在热带地区, 水体的水温变化范围较小, 也导致轮虫全年种的存在, 如果这些全年种类也是优势种或能够主导群落的总丰度或总生物量, 群落结构在时间尺度上的异质性则下降。大型水库通常划分为三个特征不同的区域: 河流区、过渡区和湖泊区, 这三个区域的群落结构通常也存在较大的空间异质性[9]。由于富营养化水平的提高会降低轮虫群落的多样性与时空异质性, 热带大型富营养化水库轮虫群落的时空异质性存在不确定性。
本文以位于广东的大型热带富营养化水库——大沙河水库为对象, 通过对代表性水域的全年逐月采样与观测, 分析河流区、过渡区和湖泊区轮虫群落结构的时空异质性, 探讨影响热带富营养水库轮虫群落结构的主要因素。
2 材料与方法
2.1 研究区域概况
大沙河水库, 是一座大型水库, 地处广东省开平市(22°32 N, 112°30 E), 位于北回归线南侧, 目前主要承担开平市 15 万居民的生活用水和周边乡镇 13.55 万亩农田的灌溉任务。水库流域面积为217 km2, 总库容 2.58×108 m3, 正常库容 1.568×108 m3; 水库有明显的河流区和湖泊区, 周边的人类活动剧烈, 是富营养化程度较高的水体[10]。
2.2 样品采集与处理
2010年1月至12月, 在大沙河水库的河流区、过渡区和湖泊区各设置了1个采样点: S1、S2、S3(图1),每月采样1次。
浮游动物定性样品用孔径为64 μm浮游生物网于水平与垂直方向拖网; 浮游动物定量样品用 5 L采水器从水面下0.5 m处开始每2 m采样直至水库底部, 共采集水样25—50 L, 用孔径38 μm的浮游生物网现场过滤浓缩。用40%的甲醛溶液对样品进行固定至甲醛终浓度5%。
同时, 在0.5 m、5 m和10 m三个水深处采集水样, 水化学分析后取平均值用于进一步分析。
2.3 水质测定与生物样品分析
水库流域内降雨量、水位、入库流量数据由水库管理处提供; pH、水温、溶解氧等用YSI水质分析仪进行了现场测定, 透明度用塞氏黑白盘测定。总磷(TP)、总氮(TN)、硝氮(NO3-)、氨氮(NH4+)、和正磷(PO43–)根据国家水质监测标准方法进行测定[11],Chla采用反复冻浸-浸提法测定[12], 每个水化样品水化指标的测定均设置三个平行, 使用平均值作为最终结果用于数据分析。本文视降雨量、水位、入库流量为水文变量, pH、溶解氧、透明度、总磷、总氮、硝氮、氨氮和正磷为水体理化变量。
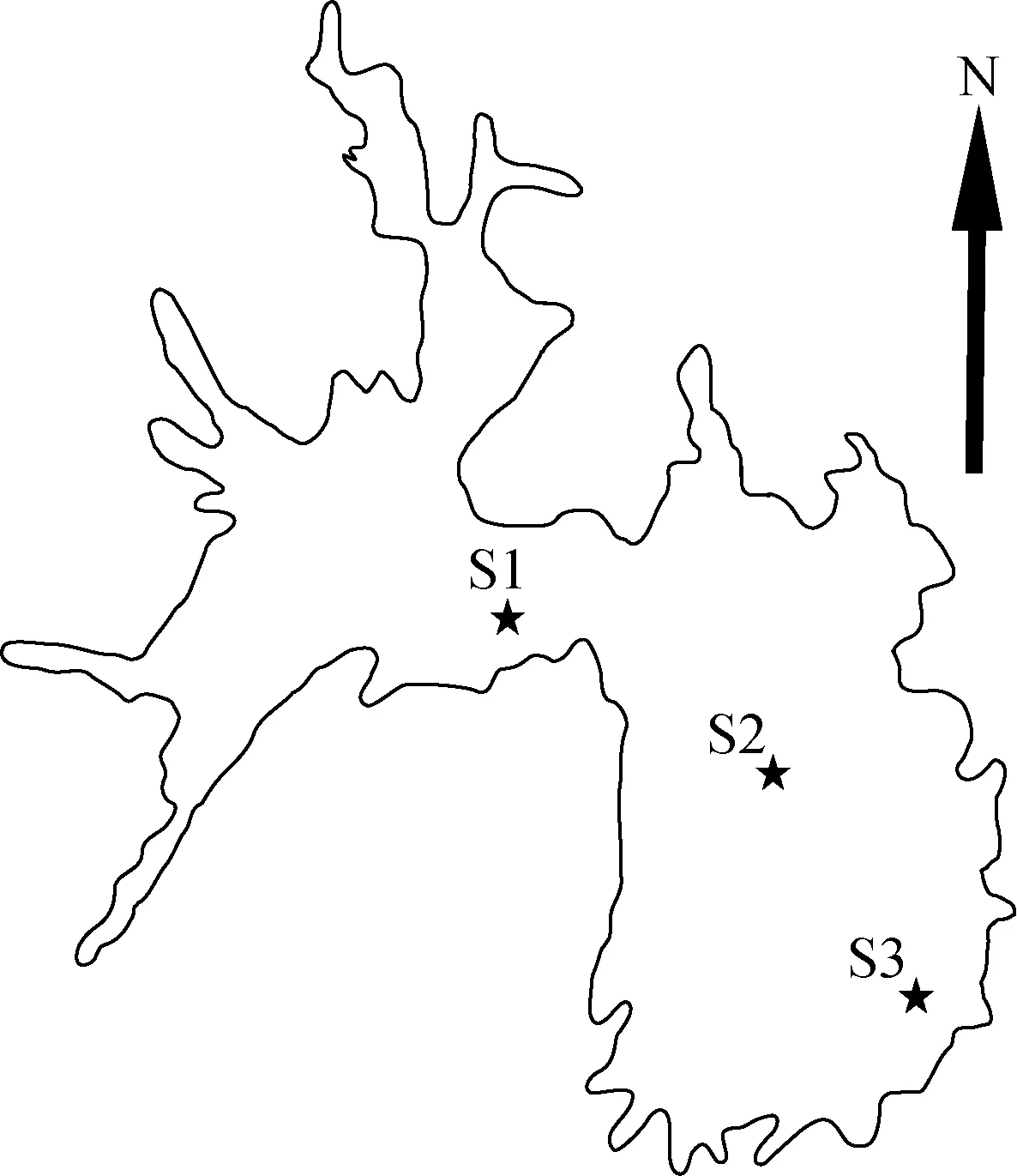
图1 大沙河水库库形和采样点的位置Fig.1 Morphology of Dashahe Reservoir and locations of three sampling sites (S1, S2 and S3)
甲壳类浮游动物在显微镜与解剖镜下进行定种和计数, 根据 Koste分类系统在显微镜下进行轮虫定种与计数。并根据浮游动物丰度计算公式计算丰度[13]。
对浮游动物计数的同时, 测量每个个体的大小。其中, 轮虫的体积通过测定体长和体宽, 采用近似几何体积公式计算体积, 使用近似密度 1 g·cm-3计算其生物量。甲壳类动物则依据体长与体重的几何异速方程计算得出每个个体的体重, 从而得到各个物种的生物量[7,13]。
2.4 数据分析
在 R语言平台上采用软件包(MASS、stats、labdsv、cluster、gclus 、vegan 和 packfor)进行浮游动物群落的多样性计算和结构比较等多元统计分析。对轮虫、枝角类和桡足类丰度数据分析前, 先用log1p转换, 再进行Hellinger转换, 以减弱双零问题带来的影响。轮虫的多样性指数采用Hill number of Shannon diversity (后用 Shannon-Hill表示); 在轮虫丰度数据对采样时间进行聚类分析时, 组间距用最小组内方差法进行计算(Ward法); 在浮游动物群落的 PCA分析中, 同样使用转换后的轮虫丰度数据; 计算物种指示值时使用 indval函数, 按聚类分析划分的季节计算不同季节的的指示物种, 选取通过置换检验(p< 0.05)的作为指示种; 对水库理化数据和轮虫物种数、生物量、丰度、多样性指数进行多因素方差分析时, 以采样点和季节作为因子变量;在RDA分析中, 解释变量由标准化后的水体理化数据和水文变量数据, 以及转化后的枝角类和桡足类丰度数据组成, 响应变量是转化后的轮虫丰度数据,RDA分析后利用前向选择剔除不显著变量[14]; 使用线性回归分析轮虫生物量与枝角类生物量和轮虫生物量与桡足类生物量, 并对回归的显著性做F检验。
3 结果与分析
3.1 水体理化特征
大沙河水库全年水温在15.40—32.30 ℃内变化,三个采样点间水温变化趋势相似, 其中 4—10月平均温度为 28.59 ℃, 1—3月平均温度 17.00 ℃,11—12月平均温度 21.07 ℃, 最高温度在 7月, 最低温度在 1, 2月(图 2a); 水库月降雨量介于0—543.80 mm之间, 全年波动较大, 4—9月降雨丰富, 分明显雨季(4—9月)和旱季(10—3月) (图 2b);水库入库流量介于2.72—632.24 m3·s-1之间, 全年波动较大, 分为丰水期(4—10月)和枯水期(11—3月)(图2c) ; 水位相对稳定, 介于32.67—33.73 m之间,全年变化最大值超过1 m, 夏季水位最低(图2d)。
三个采样点(S1, S2, S3)的SD、TN、TP和Chla变化 趋势相似(Pearson相关性检验p< 0.05)。水体透明度夏季最高, 春秋两季透明度较低(图 3a); 总氮变化范围为0.339—1.608 mg·L-1, 2—5月总氮较高, 7月总氮最低(图3b); 总磷变化范围为0.009—0.072 mg.L-1, 变化趋势与总氮类似(图3c); 叶绿素a浓度变化范围0.012—0.094 mg·L-1全年波动较大,年平均值为0.035 mg·L-1, 1月和11月Chla浓度最高(图 3d)。
3.2 轮虫群落
3.2.1 群落分析
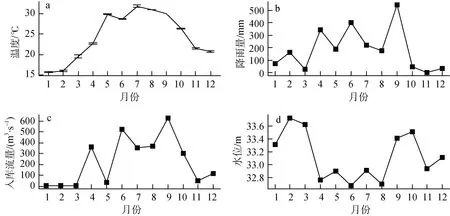
图2 大沙河水库水文变量逐月变化Fig.2 Monthly variation of the hydrological features in Dashahe reservoir
全年样品中共检出轮虫物种数25种, 19种属于游泳亚目, 4种属于簇轮亚目, 2种属于胶鞘亚目,其中全年种2种, 常见种2种, 稀有种7种(表1), 3月所有采样点的物种数都最少, 湖泊区只有3个种。夏季的物种数有增加的趋势, 3个采样点的检出的轮虫物种数具有相似的动态特征(图 4a);轮虫全年多样性指数平均值0.44, 其中7—10月最高, 平均值为 0.64, 12月多样性最低, 采样点S3的多样性在三月有一个异常高的值(图 4b); 3个采样点上轮虫群落的总丰度具有相似的变化特征, 除 12月份外, 轮虫群落的总丰度相对稳定(图4c); 3个采样点上群落生物量的平均水平接近21.131 μg·L-1, 但在夏季 3个采样点之间的有较大的差异(图4d)。

图3 大沙河水库水体理化特征Fig.3 Monthly variation of physico-chemical variables in Dashahe reservoir

表1 轮虫种类组成和出现频次Tab.1 The species of rotifers and their occurring frequency
3.2.2 群落排序和指示种
聚类分析轮虫丰度数据发现, 除S1的8月份数据外, 3个采样点的1—4月数据聚为一簇, 5—9月的数据聚为一簇, 10—12月的数据聚为一簇; 再对轮虫丰度数据进行PCA分析, 以1型标尺(scaling-1)对各个采样点的各个月份采样数据以PCA分析后的前两个主轴(累计解释率R2=0.383)做双序图(图5); 根据对各采样点的采样时间的排序结果和各月份降雨情况, 将大沙河季节划分为枯水一期(1—4月), 丰水期(5—9月)和枯水二期(10—12月), 计算各个月份的轮虫物种指示值(表2)。
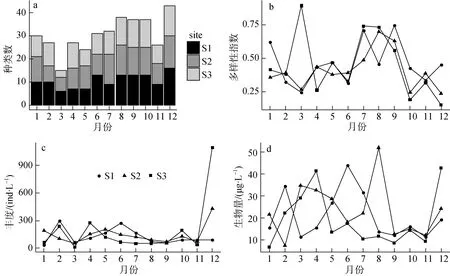
图4 轮虫种数与多样性指数(Shannon-Hill number)的逐月变化, 轮虫丰度和轮虫生物量的逐月变化规律Fig.4 Monthly variation of the richness and the Shannon-Hill number of rotifers, total abundance and total biomass ofrotifers
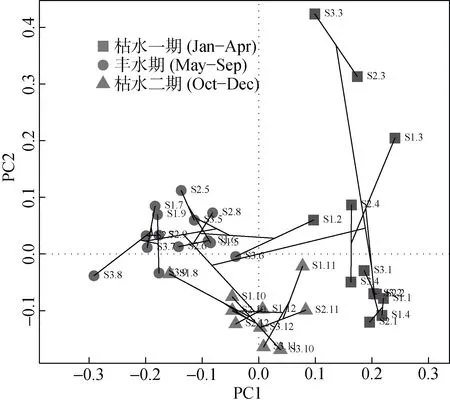
图5 轮虫丰度数据聚类与PCA联合分析。S1.1指S1采样点2010年1月的数据, S1.2指S1采样点2010年2月的数据,依次类推, 直到S3.12指S3采样点2010年12月的数据。Fig.5 PCA and cluster analysis of rotifer community in abundance.S1.1 for the data of Jan 2010 from S1, and S1.2 for the data of Feb 2010 from S2, in a similar fashion to S3.12 for the data of Dec 2010 from S3
3.3 时空异质性
3.3.1 水库理化性质的时空异质性
方差分析水库的理化变量叶绿素a浓度、总磷、总氮、温度、透明度、水位、降雨量和入库流量的结果显示, 除去降雨量、水位和入库流量是水库整体的水文变量, 不存在空间异质性的说法, 温度、总氮和叶绿素a浓度没有空间异质性(pTEM = 0.956;pTN =0.623;pChla = 0.392), 总磷和透明度具有空间异质性(pTP = 0.068;pSD = 0.047); 除叶绿素a浓度没有时间异质性外, 其余所有的变量都有时间异质性。
3.3.2 轮虫群落结构时空异质性
方差分析表明轮虫的多样性与采样点无关(p=0.898), 但是与采样的季节有关 (p= 0.004), 丰水期多样性最高, 枯水一期其次, 枯水二期多样性最低(图 6a); 不同季节样品的物种数有显著差异(p=0.007), 丰水期和枯水二期物种数较多, 枯水一期较少(图 6b); 轮虫的丰度和生物量均不具有显著的时空异质性(图6c和图6d)。

表2 群落中的指示值计算及在样品中检出频次Tab.2 Indicator species and their occurring frequency of occurrence in samples
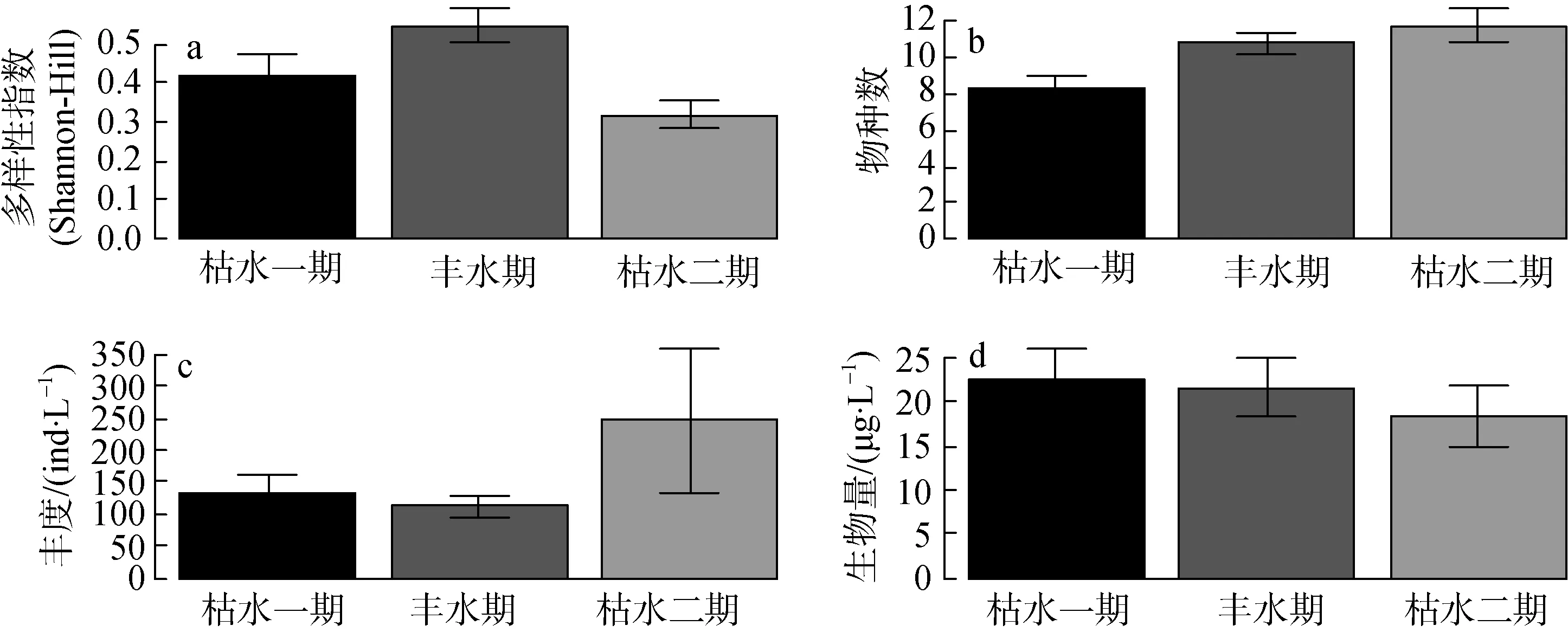
图6 轮虫群落的季节性(时间异质性)Fig.6 Temporal variation of the community structure of rotifers
3.4 轮虫丰度的RDA分析
27个解释变量中有25个变量的VIF值大于10,具极强的共线性, 前向选择后得到温度(TEM)、硝氮(NO3–)、水位(WL)、氨氮(NH4+)、正磷(PO43-)、降雨量(RAIN)、入库流量(Inflow)和叶绿素 a浓度(Chla)是对约束排序有显著贡献的解释变量(解释率由高到低), 由这些环境变量对响应变量的解释率为R2adj=0.412, 其中温度对约束排序的贡献最大为0.165, 叶绿素浓度贡献最小为0.027, 水体理化变量解释率 31.935%, 水库水文变量对轮虫群落结构变化解释率9.334%。基于2型标尺(scaling-2)三序图(图7)表明, 3个采样点的不同月份的数据排序结果和聚类分析结果相似(图5)。
3.5 轮虫、枝角类和桡足类之间的竞争与捕食
拟合轮虫多样性指数和轮虫生物量的关系, 随着轮虫生物量的增加, 轮虫多样性并没有显著变化(p=0.568); 轮虫生物量与枝角类生物量呈弱正相关(R2adj= 0.048;p= 0.105) (图8a); 桡足类生物量与轮虫生物量呈显著正相关(R2adj= 0.339;p= 0.0001) (图8b)。桡足类与轮虫生物量生物量之比的变化与轮虫生物量逐月变化趋势相似。
4 讨论与结论
大沙河水库是一座大型水库, 该水库有明显的河流区、过渡区和湖泊区, 周边的人类活动剧烈, 是富营养化程度较高的水体。大沙河水库的水文变量和理化变量, 在不同的采样点间有相似的变化趋势,同时逐月变化明显, 因此可以推测, 该水库的环境变化具有时间异质性, 但是空间异质性较小。水库叶绿素 a浓度全年波动性较大, 其变化趋势与透明度变化趋势有显著的对应关系, 叶绿素 a浓度高时,藻类丰度大, 导致水体透明度降低, 反之, 水体透明度上升; 7—10月水库来水和排水水频率增加, 导致水力滞留时间减短, 同时降水量大, 水体营养盐浓度下降, 导致藻类丰度下降, 叶绿素a浓度随之下降, 浮游动物食物数量减少。

图7 轮虫丰度数据RDA的约束排序。KCO指代螺形龟甲轮虫, KTR指代热带龟甲轮虫 (K.tropica) , CUN指代独角聚花轮虫 (C.unicorni) , TST指代对棘异尾轮虫 (T.stylata) , BCAL指代萼花臂尾轮虫 (B.calyciflorus) , TCY指代圆筒异尾轮虫(T.cylindrica) , TLO指代长刺异尾轮虫 (T.longiseta) , TRO指代罗氏异尾轮虫 (T.rousseleti) , TCA指代刺盖异尾轮虫(T.capucina) , BDO指代 (B.donne) , BFO指代剪形臂尾轮虫 (B.forficula) , BFA指代镰状臂尾轮虫 (B.falcatu) , BCAU指代尾突臂尾轮虫 (B.caudatus) , FLO指代长三肢轮虫 (F.longiseta) , FOP指代脾状三肢轮虫 (F.opoliensis) , FBR指代臂三肢轮虫 (F.brachiata) , PVU指代广生多肢轮虫 (P.vulgaris) , PEU指代真翅多肢轮虫 (P.euryptera) , HMI指代奇异六腕轮虫(H.mira) , AOV指代卵形无柄轮虫 (A.ovalis) , PSU指代沟痕泡轮虫 (P.sulcata) , APR指代前节晶囊轮虫 (A.priodonta) , PHU指代郝氏皱甲轮虫 (P.hudsoni) , LSP指代腔轮虫sp.(Lecane.sp) , COL指代胶鞘轮虫.sp (Collotheca.sp) 。Fig.7 Triplot of rotifer community in abundance with environment variables, total abundances of cladocerans and copepod used as the explaining variables in RDA.KCO- K.cochlearis, KTR-K.tropica, CUN-C.unicorni, TST- T.stylata,BCAL-B.calyciflorus, TCY-T.cylindrica, TLO-T.longiseta, TRO-T.rousseleti, TCA-T.capucina, BDO-B.donner, BFO- B.forficula,BFA-B.falcatus, BCAU-B.caudatus, FLO-F.longiseta, FOP-F.opoliensis, FBR-F.brachiata, PVU-P.vulgaris, PEU-P.euryptera,HMI-H.mira, AOV-A.ovalis, PSU- P.sulcata, APR-A.priodonta, PHU-P.hudsoni, LSP-Lecane.sp, COL- Collotheca.sp.
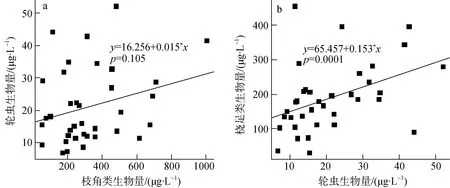
图8 轮虫生物量与枝角类生物量的关系(a)桡足类生物量与轮虫生物量的关系(b)Fig.8 Relationship between the biomass of rotifers and cladocerans (a), and the biomass of copepods and rotifers (b)
轮虫物种数全年波动较大, 在枯水一期物种数最少, 丰水期物种数最多, 同时轮虫丰度的变化趋势和轮虫生物量变化趋势差异较大, 说明轮虫的种类组成也有较明显的动态变化。一般认为, 尖尾疣毛轮虫 (Synchaeta stylata) 、郝氏皱甲轮虫、柱足腹尾轮虫 (Gastropus stylifer) 、卵形无柄轮虫和独角聚花轮虫是寡营养水体的优势种; 中营养水体种类较多, 一般为过渡种; 臂尾轮虫、裂痕龟纹轮虫(Anuraeopsis fissa) 、沟痕泡轮虫、异尾轮虫、长三肢轮虫、奇异六腕轮虫、多肢轮虫和螺形龟甲轮虫等是富营养水体的优势种[15]。在大沙河水库中, 螺形龟甲轮虫、异尾轮虫, 沟痕泡轮虫和多肢轮虫丰度全年较高, 因此, 从轮虫种类组成上看, 大沙河水库以富营养化种类为主, 但独角聚花轮虫在轮虫全年都是优势种, 可能是因为以群聚的方式生长, 因而受到的捕食压力相对与其他躲避捕食者能力较差的轮虫较小[16], 在捕食压力下具有一定的竞争优势。
轮虫和枝角类的生长都较为迅速, 枝角类群落结构更多的受到下行效应的影响, 然而轮虫更多的受到上行效应影响, 对水体理化环境变化响应迅速,因此许多研究都倾向于以轮虫作为指示种去指示水体的变化[17]。对处于富营养型的大沙河而言, 指示种可以对浮游群落结构变化具有很好的指示意义[18]。利用 indval计算得到大沙河各个季节的指示种(表2),对比轮虫在各个月份出现的情况, 单一物种不具有很好的指示意义, 需要针对某个季节的多个指示种同时出现时, 指示种才有意义。我们结合RDA三序图: 枯水一期指示种为独角聚花轮虫、真翅多肢轮虫和罗氏异尾轮虫, 这几种物种的出现与高硝氮浓度和高氨氮浓度相关; 丰水期指示种为对棘异尾轮虫、奇异六腕轮虫和剪形臂尾轮虫, 与高温、高降雨量和大的入库流量相关; 枯水二期指示种为广生多肢轮虫、胶鞘轮虫、卵形无柄轮虫和长刺异尾轮虫与高正磷浓度相关。可以很明显看到, 枯水一期和枯水二期与指示种相关的环境变量分别是氮和磷,而因此可以推测, 枯水一期和枯水二期, 轮虫群落结构的变化可能与氮限制和磷限制浮游植物的大量繁殖有关, 而丰水期轮虫群落结构的变化可能与轮虫对温度的响应差异和轮虫的游泳能力差异有关。
从聚类分析的结果来看, 大沙河的季节变化属于典型的南亚热带季风气候, 全年可分为枯水一期(1—4月), 丰水期(5—9月)和枯水二期(10—12月)。以聚类分析的季节划分方式作和采样点作为因子的方差分析显示, 大沙河水库的理化环境具有明显的时间异质性, 空间异质性不明显, 轮虫物种数和多样性指数亦具有时间异质性, 呈现出明显的季节划分, 枯水一期物种数最少, 丰水期物种数最多, 同样轮虫的群落结构也不具有空间异质性。春季气温回暖, 藻类大量繁殖, 为轮虫、枝角类和桡足类提供了充足的食物, 大型枝角类受鱼类的捕食压力极高,只有小型枝角类、桡足类和轮虫生长[2,4,19], 随着浮游动物数的增加, 枝角类和轮虫的竞争与桡足类对轮虫的捕食加强, 食物补给不足, 从而使轮虫丰度在春季中期开始降低; 丰水期由于降水量大, 以及为了防洪换水频率增加, 水力滞留时间显著缩短,一方面使藻类密度降低, 另一方面换水过程也使得轮虫个体损失较多, 同时水体透明度增加, 使枝角类和桡足类受到更大的鱼类捕食压力, 降低轮虫的竞争压力和捕食压力, 轮虫物种数和多样性保持一个较高的水平[20–21]; 枯水二期轮虫群落结构变化大致同春季相同, 但是在2010年11月发生蓝藻水华,12月水华消退后, 水体有机质和含量大幅增加, 为轮虫提供了丰富的食物来源, 因而使得轮虫多样性指数、生物量和丰度异常的高, 也使得12月份指示种分析出现异常。
冗余度分析(RDA)显示影响轮虫群落结构的主要是水库水体的理化变量, 包括温度、硝氮、氨氮、正磷和叶绿素 a浓度, 它们共解释了轮虫群落结构变异的31.935%, 而水文变量——水位、降雨量和入库流量, 一共解释了 9.334%。受季风影响, 水温通过影响轮虫的生长发育, 浮游甲壳类动物的生长和食物(叶绿素a和温度显著负相关, 线性回归R2adj=0.192,p= 0.004)的供给而显著改变轮虫的群落结构[7,22]; 水库水位全年变化较大, 最大变幅为 1.06m,植物沿岸带难以维持, 水体生境多样性降低, 降低了轮虫群落结构的空间异质性[23], 但空间异质性较低也可能是因为在三个采样水域空间差异小, 大沙河水库地势平坦, 水库生态分区不明显[10], 同时水体富营养化也一定程度上降低了轮虫种类组成的多样性, 从而进一步降低了轮虫群落结构的空间异质性[24]; 叶绿素 a浓度可以一部分代表轮虫的食物来源, 虽然叶绿素a是显著的解释变量, 但是其解释率最低, 这可能是因为叶绿素a浓度变化是对温度、降雨和营养盐等其他变量变化的响应, 与其他解释变量相关。
轮虫多样性指数和轮虫生物量关系不显著, 反映在捕食压力下, 少数轮虫难以形成绝对优势种,轮虫种间竞争较弱。轮虫生物量与枝角类生物量呈弱正相关, 显示轮虫与枝角类的竞争较弱; 桡足类的生物量与轮虫生物量显著正相关, 可能是桡足类对轮虫有较强捕食作用的结果, 这与桡足类种类组成以温中剑水蚤 (Mesocyclops thermocyclopoides)这样的捕食性桡足类为主的特征相符合[25–27]。在RDA前向选择之前, 桡足类和枝角类丰度作为解释变量, 具有极高的膨胀系数, 前向选择后桡足类和枝角类都被排除, 因此我们可以推测, 桡足类对轮虫捕食作用的动态变化与水库水文状况和理化特征的变化是相关的。
大沙河水库中轮虫的群落具有明显的枯水一期、丰水期和枯水二期 3个时期更替, 轮虫多样性丰水期最高, 枯水期多样性较低, 群落结构变化受温度和入库流量等水体理化和水库水文变量的影响,不同季节具各自的指示种; 受鱼类等捕食者影响,枝角类与轮虫之间的竞争强度较低, 但桡足类对轮虫可能有显著的捕食作用, 轮虫种间竞争不显著;由于水库富营养化程度高, 三个水域的环境变量相对一致, 轮虫群落结构没有明显的空间异质性。
[1]杨柳, 陈绵润, 林秋奇, 等, 一座热带高产渔业水库枯水期轮虫的群落组成与动态分析[J].湖泊科学, 2008, 20(6):780–789.
[2]GILBERT J J.Competition between rotifers and Daphnia[J].Ecology, 1985, 66(6): 1943–1950.
[3]GILBERT J J.Suppression of rotifer populations by Daphnia: A review of the evidence, the mechanisms, and the effects on zooplankton community structure1[J].Limnology and Oceanography, 1988, 33(6): 1286–1303.
[4]MACISAAC H J, GILBERT J J.Competition between rotifers and cladocerans of different body sizes[J].Oecologia, 1989, 81(3): 295–301.
[5]DODSON S I, ARNOTT S E, COTTINGHAM K L.The relationship in lake communities between primary productivity and species richness[J].Ecology, 2000, 81(10):2662–2679.
[6]PROULX M, PICK F R, MAZUMDER A, et al.Experimental evidence for interactive impacts of human activities on lake algal species richness[J].Oikos, 1996:191-195.
[7]林秋奇.流溪河水库后生浮游动物多样性与群落结构的时空异质性[D].广州: 暨南大学, 2007.
[8]秦伯强, 高光, 朱广伟, 等.湖泊富营养化及其生态系统响应[J].科学通报, 2013, 58(10): 855-864.
[9]BETSILL R K, van den Avyle M J.Spatial heterogeneity of reservoir zooplankton: a matter of timing?[J].Hydrobiologia, 1994, 277(1): 63–70.
[10]郭菲菲, 李慧明, 顾杨亮, 等.大沙河水库浮游甲壳类动物群落特征[J].生态科学, 2015, 34(5): 8–15.
[11]王心芳, 魏复盛, 齐文启.水和废水监测分析方法[M].北京: 中国环境出版社, 2002: 216–219.
[12]林少君, 贺立静, 黄沛生, 等.浮游植物中叶绿素a提取方法的比较与改进[J].生态科学, 2005, 24(1): 9–11.
[13]章宗涉.黄祥飞.淡水浮游生物研究方法[M].北京: 科学出版社, 1991.
[14]BORCARD D, GILLET F, LEGENDRE P.Numerical ecology with R[M].Springer Science & Business Media,2011.
[15]王晓辉, 望甜, 林秋奇, 等.南亚热带富营养化抽水型水库轮虫的组成与动态[J].湖泊科学, 2009, 21(1):101–109.
[16]王家辑.中国淡水轮虫志[M].北京: 科学出版社, 1961.
[17]BRANCO, C.W., ROCHA, M.I.A., PINTO, G.F., et al.Limnological features of Funil Reservoir (RJ, Brazil) and indicator properties of rotifers and cladocerans of the zooplankton community.Lakes & Reservoirs: Research &Management, 2002, 7(2): 87–92.
[18]GANNON J E, STEMBERGER R S.Zooplankton(especially crustaceans and rotifers) as indicators of water quality[J].Transactions of the American Microscopical Society, 1978, 97(1): 16–35.
[19]BROOKS J L, DODSON S I.Predation, body size, and composition of plankton[J].Science, 1965, 150(3692):28–35.
[20]LIN Qiuqi, DUAN Shunshan, HU Ren, et al.Zooplankton distribution in tropical reservoirs, South China.International Review of Hydrobiology, 2003, 88(6):602–613.
[21]林秋奇,赵帅营,韩博平.广东省水库轮虫分布特征[J].生态学报, 2005, 25(5): 1123–1131.
[22]ALLAN J D.Life history patterns in zooplankton[J].The American Naturalist, 1976, 110(971): 165–180.
[23]SCHRIVER, P.E.R., BØGESTRAND, J., JEPPESEN, E.,et al.Impact of submerged macrophytes on fish‐zooplanl phytoplankton interactions: large‐scale enclosure experiments in a shallow eutrophic lake.Freshwater Biology, 1995, 33(2): 255–270.
[24]钱方平,席贻龙,温新利等.湖泊富营养化对轮虫群落结构及物种多样性的影响[J].生物多样性, 2007, 15(4):344–355.
[25]BRANDL Z.Freshwater copepods and rotifers: predators and their prey[J].Hydrobiologia, 2005, 546(1): 475–489.
[26]KUMAR R, RAO T R.Effect of the cyclopoid copepod Mesocyclops thermocyclopoides on the interactions between the predatory rotifer Asplanchna intermedia and its prey Brachionus calyciflorus and B.angularis[J].Hydrobiologia, 2001, 453(1): 261–268.
[27]RAO T R, KUMAR R.Patterns of prey selectivity in the cyclopoid copepod Mesocyclops thermocyclopoides[J].Aquatic Ecology, 2002, 36(3): 411–424.
Temporal and spatial variation of the community structure of rotifers in Dashahe Reservoir, southern China (Guangdong province)
XU Shaolin, LI Huiming, GU Yangliang, HAN Boping*
Faculty of Ecology,Jinan University,Guangzhou510632,China
To investigate the temporal and spatial variation of rotifer community structure in tropical large eutrophication reservoirs, 36 samples in riverine, transition and lacustrine zones of the Dashahe Reservoir (Guangdong province, China)were gathered monthly in 2010; physic-chemical variables and zooplankton community data were also collected.25 species were identified.Among them, 19 species belonged to 6 families, 8 genera of Ploima, and 4 species from 1 family, 2 genera of Flosculariacea and the remaining 2 species belonged to 2 families, 2 genera of Collothecacea.The results are as follows.(1)There existed significantly difference among different seasons; the time series samples could be divided into three season clusters: Dry-season-I(Jan-Apr), Flood-season(Mar-Sep), Dry-season-II(Oct-Dec).Indval analysis showed there were no less than three different rotifers seen as indicator species for each season.(2) Spatial variation of the community structure and biodiversity of rotifers (Shannon-Hill number) was not significant, but there was common seasonality of the diversity and richness of rotifers (p= 0.004,p=0.007) among the three temporal group.(3) 41.269% of all the variation of the community structure of rotifers was explained by environmental variables; physic-chemical variables which included water temperature variable (explained 16.512%) explained 31.935% of all those variation; and hydrological variables explained 9.334% of the rest, which showed the nonnegligible importance of physic-chemical variable of reservoirs to rotifer community structure.
rotifers; community structure; temporal variation; spatial variation; hydrology
10.14108/j.cnki.1008-8873.2017.05.003
Q145
A
1008-8873(2017)05-018-09
徐少林, 李慧明, 顾杨亮, 等.广东省大沙河水库轮虫群落结构的时空异质性分析[J].生态科学, 2017, 36(5): 18-26.
XU Shaolin, LI Huiming, GU Yangliang, et al.Temporal and spatial variation of the community structure of rotifers in Dashahe Reservoir, southern China (Guangdong province)[J].Ecological Science, 2017, 36(5): 18-26.
2016-08-26;
2016-10-23
广东省水资源节约与保护专项资金(201303); 国家自然科学基金项目(31170436)
徐少林(1993—), 男, 2016级在读研究生, 从事数量生态学、种群遗传学, E-mail: 15521332622@163.com
*通信作者:韩博平, 教授, E-mail: tbphan@jnu.edu.cn
