水稻脱粒性的研究进展及展望
2017-08-22王继恩邱东峰张再君
王继恩+邱东峰+张再君
摘要:水稻是重要的粮食作物,提高水稻产量是水稻育种的重要目标。在水稻驯化中,许多性状朝着有利于增产的方向发展,尤其是水稻脱粒性的变化。相对于野生稻而言,栽培稻倾向于难脱粒的表型。这有效地保证了水稻谷粒的收获,进而提高了水稻产量。为了更好地理解水稻脱粒性,就水稻脱粒的细胞学基础,基因调控机制,植物激素调节做了相关综述,以期为水稻脱粒性的研究提供有用的信息,并在此基础之上对其做了一些展望。
关键词:脱粒性;离层;木质素
中图分类号:S511 文献标识码:A 文章编号:0439-8114(2017)14-2601-04
DOI:10.14088/j.cnki.issn0439-8114.2017.14.001
Abstract: Rice is an important crop, and improving rice yield is an important goal of rice breeding. In the domestication of rice, many traits in favor of yield direction. Especially for rice shattering, compared with wild rice,cultivated rice tends to be difficult to shattering phenotype. This effectively ensured the harvest of rice grain,and thus increased the yield of rice. In order to understand rice shattering better,this paper reviewed the cytological basis,gene regulation mechanism and plant hormone regulation of rice shattering. so as to provide useful information for rice shattering study. And on the basis of which we have done some discussion and outlook.
Key words: shattering; abscission layer; lignin
水稻作为重要的粮食作物,其产量的提高关乎民生大计。矮化品种(IR8)和杂交水稻(两系/三系)的培育使得水稻产量有了很大提高[1]。近年来基于籼粳杂交,提出了通过分子模块设计,结合理想株型开发高产、优质、抗逆水稻的新途径。这对于水稻产量的进一步提高具有重要意义[2]。
考古学证据表明在一万年前,我们的祖先就开始了对野生稻的驯化[3]。在驯化中许多性状(株型、穗型、粒型、脱粒性)朝着有利于水稻产量提升的方向进化着。特别是水稻脱粒性的变化,对于野生稻而言,成熟谷粒趋向于易脱落,这对于种群的繁衍具有重要的意义。然而在农业生产中,这种易脱粒性状却给栽培稻的收获带来了很大不便,它导致了收获指数(水稻实际结实数与人工收获到的谷粒数比值)下降,进而影响了水稻产量[4]。研究表明,不脱粒表型出现在约6 600年前[5]。而在栽培稻中,粳稻更倾向于难脱粒。因此在脱粒性状的研究中,多以粳稻(日本晴)作为难脱粒材料。在实际生产中,收割器械对水稻的收获产量也有影响,无论是易脱粒品种还是难脱粒品种都不能被有效收获,因此筛选及培育中等落粒程度水稻品种对水稻产量的提高具有重要意义[6]。
目前对于水稻脱粒性评估方法主要有下落法、揉搓法以及仪器测定法。这三种方法各有优劣。但目前仪器测定法是对水稻脱粒性评估比较有效的方案。利用数字测力计检测谷粒BTS(Breaking tensile strength)值来对水稻脱粒性进行评价,因为BTS值与脱粒程度的高低呈负相关,即BTS值越小,水稻谷粒越容易脱落[7]。为了加深对水稻脱粒性的研究,本研究综述了水稻脱粒的细胞学基础、基因调控机制、植物激素调节,以期为水稻脱粒性的研究提供有用的信息,并在此基础之上对其做了一些展望。
1 水稻脫粒性的细胞学基础
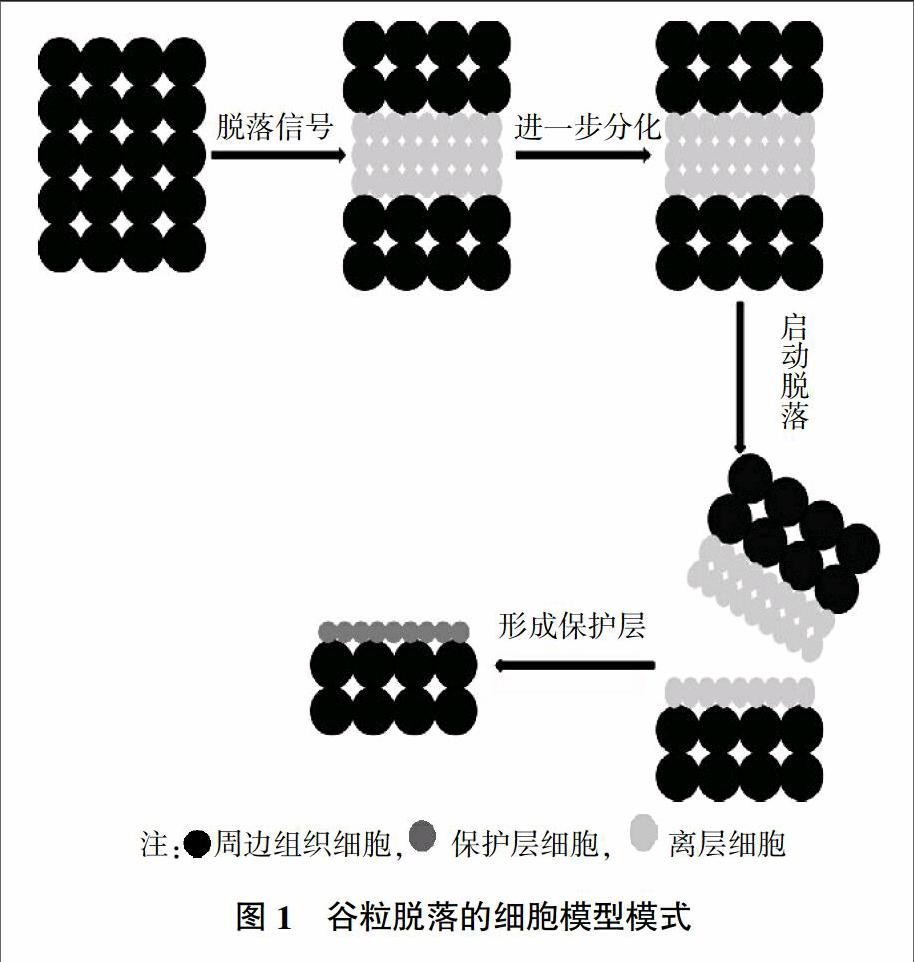
在自然界中,脱落(Abscission)是一种普遍存在的现象。一般而言脱落是对组织或器官,由于自身或环境(冷/旱胁迫)因素,与母体(或主体)发生分离的综合描述。如种子脱落、落叶、落果。这些事件的发生涉及特定细胞发育的调控。对于水稻而言,研究表明其谷粒的脱落与水稻护颖与枝梗之间离层(Abscission Layer)的发育调控有关。从水稻生命周期来看,造成水稻谷粒脱落的离层在水稻抽穗前16~20 d就开始发育,即幼穗中配子细胞起始分化的阶段。电镜组织切片观察,在水稻护颖和枝梗之间的离层是由几层小且致密的薄壁细胞组成,在抽穗期其细胞群扩展到最大[8]。从细胞形态上来看,发现这部分离区细胞明显区别于周围细胞,因此关于此处细胞形态的建成对谷粒脱落至关重要。对拟南芥研究发现TCS1在细胞形态塑造中发挥着重要作用,根据趋同进化,推测在水稻离区可能也存在类似TCS1的调控机制来影响离区细胞分化,进而影响水稻脱粒[9]。
在栽培稻中,粳稻中离区的分化要晚于籼稻,这可能与籼稻的生长周期短于粳稻有关。而在某些粳稻中没有发现离区的形成,这类粳稻通常表现为难脱粒的表型。在拟南芥及禾本科植物(水稻、高粱)中的研究表明,在脱落起始的部位,一些细胞接受环境或内源信号诱导,启动离层细胞分化,在特定的时间启动脱落信号,激活相关基因,引发谷粒脱落。这一系列的通路中涉及了细胞形态的改变及水解酶的释放(破坏细胞层之间的胞间层)。最近的研究表明,脱粒性可能涉及离层细胞的木质化,被认为是一种程序性细胞凋亡。
无论从形态学还是解剖学角度,均能清晰的将离层细胞与周围组织细胞区分开来。这群小而致密的薄壁细胞通过胞间连丝相连,形成层状的细胞群。到目前为止,关于谷粒脱落的细胞模型已基本成型(图1),主要分为以下四个阶段:①细胞在分离区分化形成参与脱落的离层细胞;②离层细胞响应各种细胞脱落信号;③在离层细胞内激活脱落程序;④分离区保护细胞的形成。在整个脱落过程中受很多发育和环境因素的影响。如末端器官感知到胁迫(旱/冷),开始衰老产生脱落信号,传递到离层细胞中诱发脱落[10]。
2 水稻脱粒性的基因调控
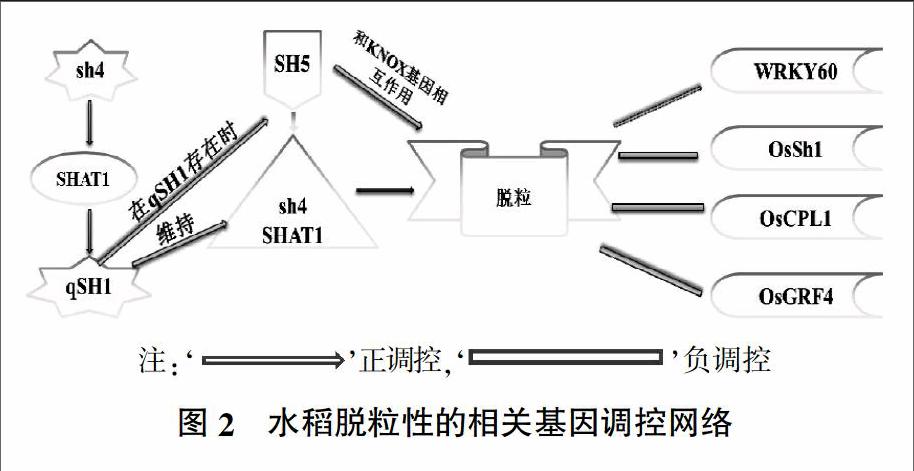
为了更好的理解水稻脱粒的调控机制,许多科学家致力于探索水稻脱粒的基因网络调控,并取得了一些重要成果[7]。目前已经克隆了若干调控水稻脱粒的基因(如qSH1、sh4、SHAT1),并对其调控机制做了进一步阐释(图2)。除此之外还发现许多其他QTLs(Quantitative trait locus,数量性状基因座)可能与脱粒有关[1]。事实上在栽培稻中水稻谷粒的脱落程度也是多样化的,这也在一定程度上说明水稻脱粒性是受多基因调控的性状[11]。下面对一些已克隆的调控基因做简单介绍。
最早克隆的两个主效基因qSH1和sh4,分别位于1号和4号染色体。qSH1和sh4分别解释了68.6%和69.0%的表型变异。研究表明,qSH1编码一个BEL1同源域转录因子,在其5′调控区一个SNP(G/T)导致其表达缺陷,影响了离层细胞的分化,表现为脱粒程度的变化[4,12]。sh4编码一个Trihelix家族(植物特有)转录因子[13]。核苷酸序列比对发现237位的SNP(G/T),导致了第79位的赖氨酸替换成天冬酰胺,该氨基酸位于Trihelix家族蛋白的重要功能域上,进而造成蛋白功能改变,影响了离层的发育,同样改变了水稻脱粒程度[14]。后续的研究中发现sh4第二外显子的单个核苷酸插入导致移码突变,产生无效等位基因,导致离层发育缺陷。以上研究表明水稻脱粒性与离层细胞的发育关系密切。
利用诱导突变系克隆了SHAT1,也位于4号染色体,该基因编码AP2(Apetala2)蛋白[8]。RNAi试验结果表明,SHAT1的低表达导致水稻脱粒性降低,并发现其与水稻离区的发育有关。研究还发现SHAT1受sh4的正调控,而SHAT1调控下游qSH1的表达。qSH1通过反馈维持SHAT1和sh4在离区细胞的表达,三者协同调控水稻脱粒性[15]。
OsCPL1是一個磷酸酶,编码C-末端结构域磷酸酶样蛋白,定位在细胞核中。研究表明,水稻中OsCPL1的失活加强了谷粒脱落,并且离层细胞得以分化。进一步研究表明,OsCPL1通过影响RNAPII(RNA聚合酶II)的CTD结构域的去磷酸化,调节下游靶基因的转录表达[16]。OsGRF4作为生长调节因子,是在2号染色体克隆的第一个与水稻脱粒有关的基因。该蛋白有两个重要结构域,WRC和QLQ,分别负责结合DNA和蛋白质。研究表明,OsGRF4是一个多效基因,其正调控粒型及穗型,负调控脱粒。在水稻发育中OsGRF4受到miR396的负调控,因此可以通过对“miR396-OsGRF4”分子模块的精妙设计来降低水稻脱粒,提高水稻产量[2,17]。OsSh1和WRKY是两个根据高粱中的同源基因发现的影响水稻脱粒性的基因,前者与离层有关,后者涉及木质素调控途径,验证表明二者均与水稻脱粒性有关[6,18]。
SH5是最近发现的与水稻脱粒性有关的基因。该基因位于5号染色体,编码BEL1蛋白,与qSH1高度同源。而BELL家族蛋白与KNOX家族蛋白相互作用。有研究表明,KNOX(Knotted-like homeobox)与BELL家族相互作用来调节下游靶基因的表达,进而调控分生组织中激素的动态平衡。因此推测SH5也可能与KNOX蛋白作用调节离层的发育,影响种子脱落。对拟南芥的研究表明BP(Brevipedicellus,KNOX家族蛋白)通过调节木质素的生物合成途径来调控细胞分化。而对于水稻中SH5是否与OSH1和OSH15(BP同源基因)相互作用还有待进一步研究。SH5在植物许多部位(根、茎、叶、穗)均有表达,但在花梗的离区表达最强。SH5过表达系中SHAT1和SH4均上调表达,表明二者均受到SH5的诱导,并且SH5需要在qSH1辅助下调节离层发育[19]。综上所述,水稻脱粒性是一个复杂的数量性状,涉及多基因调控。其完整的基因调控网络还有待进一步探索。
3 水稻脱粒的相关激素调节
直系同源基因的共线进化也促进了水稻脱粒性的研究。即不同物种中因同源基因的变化而造成相似的表型[5]。水稻护颖与枝梗之间离层对谷粒脱落至关重要,并且受很多因素的影响[6]。研究表明,IND(Indehiscent)维持离区细胞中生长素的低浓度,可以调节拟南芥果荚爆裂,种子脱落[20]。虽然目前在水稻中还没鉴定出对应的同源基因,但这也在一定程度上说明了生长素对种子脱落有影响。对OsGRF4的研究发现其调控CKX5和CKX1(细胞分裂素脱氢酶前体基因)的活性,影响了细胞分裂素的动态平衡,进而表现为穗型及脱粒性的改变[17]。这说明细胞分裂素也参与了水稻的脱粒性。前面已经知道脱粒相关基因SH5通过与KNOX蛋白相互作用,影响水稻脱粒。在玉米分生组织中KN1(KNOX家族蛋白)诱导GA2ox1表达,在水稻花序分生组织OsGA3ox2,SLR1,OsGA20ox2也是高表达的。因此推测赤霉素在水稻脱粒中可能扮演着重要角色。虽然乙烯参与果实脱落(如苹果),但其在水稻脱粒中的作用还有待进一步研究[21]。另外值得注意的是SH5和WRKY60都涉及了木质素的代谢调控,这可能对水稻脱粒性的研究提供了又一重要方向[18,19]。
4 讨论与展望
脱粒性是作物驯化的一个重要的农艺学性状,脱粒程度的高低是影响作物收获产量的一个重要指标。事实上,在水稻中脱粒性作为重要农艺性状影响了水稻产量,是一种典型的植物器官脱落现象。研究表明,器官的脱落常常伴随离区细胞的细胞间质和细胞壁的降解,进而导致器官(如种子、叶、花)脱离母体(即脱落现象)[22]。离区的发育与功能行使是多基因参与的复杂精细的调控过程,且脱粒程度多样化,使得水稻脱粒的研究进展缓慢。目前对水稻脱粒性的研究主要集中在对水稻护颖及枝梗之间的离层的发育调控上,且关注在离层发育调控的早期,即离层起始发育的时期。研究表明,离层的分化与脱粒的难易程度显著相关[6]。然而对于离层细胞发育后期的调控以及响应激素调节的分子机制尚不明朗,还有待进一步的研究。而在对基因组重测序中发现几个参与激素代谢的基因位点,可能影响了水稻脱粒。如gibberellin 20 oxidase 2(LOC_Os01g66100)参与赤霉素代谢;LOC_Os09g08130预测参与生长素诱导的磷酸化。
另外環境因素也对水稻脱粒产生了重要影响。例如研究表明冷胁迫诱导了相关脱粒基因的表达以及影响了离区的分化[23]。正如前面所述,种子脱落脱离母体,意味着其与母体细胞的之间的“连接”不存在了,也就是说发生了细胞“消失”[10]。在一些情况下,这些过程可以由PCD(细胞凋亡)来完成。研究表明,在拟南芥花器官,成熟果实的离区观察到PCD标记BFN1和PCD相关DUF679-结构域膜蛋白(DMP)同源基因的表达,并且番茄器官的脱落也与BFN1同源物和核糖体酶(LX)的表达有关。这表明这些过程均涉及PCD途径。而相关研究在水稻中还未曾报道,并且在拟南芥中过表达PCS1引起花药DZ(离区)关闭PCD过程,进而造成不爆荚。在已获得的重测序注释位点中发现了可能参与细胞凋亡的变异位点。如LOC_Os10g17950和LOC_Os05g 45000,前者拥有PCD相关的DUF样结构域,可能参与细胞凋亡;后者为衰老诱导性的受体样蛋白激酶。目前在水稻脱粒性研究,细胞凋亡的探讨还存在很多不足,因此加强此通路的研究,对完善水稻脱粒性调控网络意义重大[24]。综上所述,水稻脱粒性是一种重要且复杂的综合性状,涉及众多因素的影响,进一步探索脱粒性相关基因的互作及与环境的关系,对深入理解水稻驯化具有重要意义。
水稻基因OsGRF4高表达导致大粒、长穗、及降低谷粒脱落。研究表明,在谷粒发育中,OsGRF4受miR396负调控[2]。这说明miRNA参与了水稻脱粒性的调控。另外在拟南芥中研究发现AP2结构域可以与miRNA172相互作用,进而抑制AP2基因的表达[25]。在水稻中SHAT1含有AP2结构域,在水稻中是否也存在类似的miRNA调控水稻脱粒还有待进一步的研究。在全基因组重测序中发现若干ncRNA,可能参与类似的调控机制,来影响脱粒相关基因的表达,进而影响水稻脱粒表型的改变。
目前对水稻脱粒性状的研究主要借助QTLs定位(sh4/qSH1)、突变体筛选(SHAT1)、同源克隆(WRKY60/OsSh1)。尤其是近年来对水稻基因组测序的完成,大大促进了水稻脱粒性的研究,并从全基因组的角度探讨了水稻脱粒性驯化的机制。本实验室利用难脱粒水稻品种及其易脱粒突变体进行全基因组重测序,结合生信分析与文献报道筛选到若干变异位点,以便进一步的试验研究。电镜技术(电镜观察)和转录组/蛋白组也将促进科研人员更快的解析调控水稻脱粒的分子机制。如2015年日本科学家利用Mutmap方法结合转录组学,仅用两年时间就培育了Kaijin耐盐品种并推广播种[26]。这同样适用于水稻脱粒性的研究。在农业生产中,实现高效的收获,规避不必要的产量损失,为粮食安全做出一定贡献。
参考文献:
[1] CHENG J,HE Y,ZHAN C,et al. Identification and characterization of quantitative trait loci for shattering in rice landrace jiucaiqing from Taihu lake valley,China[J].The Plant Genome,2016,9(3):1-9.
[2] WANG S,LI S,LIU Q,et al. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J].Nature genetics,2015,47(8):949-954.
[3] CAI H W,MORISHIMA H. Genomic regions affecting seed shattering and seed dormancy in rice[J].Theoretical and Applied Genetics,2000,100(6):840-846.
[4] KONISHI S,IZAWA T,LIN S Y,et al. An SNP caused loss of seed shattering during rice domestication[J]. Science,2006,312(5778):1392-1396.
[5] LI L F,OLSEN K M. To Have and to hold:Selection for seed and fruit retention during crop domestication[J].Current Topics in Developmental Biology,2016,119:63-109.
[6] LIN Z,LI X,SHANNON L M,et al. Parallel domestication of the Shattering1 genes in cereals[J]. Nature genetics,2012,44(6):720-724.
[7] QI X,LIU Y,VIGUEIRA C C,et al. More than one way to evolve a weed:Parallel evolution of US weedy rice through independent genetic mechanisms[J].Molecular Ecology,2015,24(13):3329-3344.
[8] YAO N,WANG L,YAN H,et al. Mapping quantitative trait loci (QTL) determining seed-shattering in weedy rice: Evolution of seed shattering in weedy rice through de-domestication[J]. Euphytica,2015,204(3):513-522.
[9] ZHOU Y,LU D,LI C,et al. Genetic control of seed shattering in rice by the APETALA2 transcription factor SHATTERING ABORTION1[J].The Plant Cell,2012,24(3):1034-1048.
[10] ESTORNELL L H,AGUST?魱 J,MERELO P,et al. Elucidating mechanisms underlying organ abscission[J].Plant Science,2013, 199:48-60.
[11] LIN Z,GRIFFITH M E,LI X,et al. Origin of seed shattering in rice(Oryza sativa L.)[J]. Planta,2007,226(1):11-20.
[12] MAGWA R A,ZHAO H,YAO W,et al. Genome-wide association analysisfor awn length linked to the seed shattering gene qSH1 in rice[J].Journal of Genetics,2016,95(3):639-646.
[13] YAN H,MA L,WANG Z,et al. Multiple tissue-specific expression of rice seed-shattering gene SH4 regulated by its promoter pSH4[J].Rice,2015,8(1):1.
[14] LI C,ZHOU A,SANG T. Rice domestication by reducing shattering[J].Science,2006,311(5769):1936-1939.
[15] DONG Y,WANG Y Z. Seed shattering:From models to crops[J].Frontiers in Plant Science,2015,6:1-13.
[16] JI H,KIM S R,KIM Y H,et al. Inactivation of the CTD phosphatase-like gene OsCPL1 enhances the development of the abscission layer and seed shattering in rice[J].The Plant Journal,2010,61(1):96-106.
[17] SUN P,ZHANG W,WANG Y,et al. OsGRF4 controls grain shape,panicle length and seed shattering in rice[J].Journal of Integrative Plant Biology,2016,58(10):836-847.
[18] TANG H,CUEVAS H E,DAS S,et al. Seed shattering in a wild sorghum is conferred by a locus unrelated to domestication[J].Proceedings of the National Academy of Sciences,2013, 110(39):15824-15829.
[19] YOON J,CHO L H,KIM S L,et al. The BEL1-type homeobox gene SH5 induces seed shattering by enhancing abscission-zone development and inhibiting lignin biosynthesis[J].The Plant Journal,2014,79(5):717-728.
[20] SOREFAN K,GIRIN T,LILJEGREN S J,et al. A regulated auxin minimum is required for seed dispersal in Arabidopsis[J].Nature,2009,459(7246):583-586.
[21] DAL C V,DANESIN M,BOSCHETTI A,et al. Ethylene biosynthesis and perception in apple fruitlet abscission(Malus domestica L. Borck)[J].Journal of Experimental Botany,2005, 56(421):2995-3005.
[22] 王 翔,陳晓博,李爱丽,等.植物器官脱落分子生物学研究进展[J].作物学报,2009,35(3):381-387.
[23] SUBUDHI P K,SINGH P K,DELEON T,et al. Mapping of seed shattering loci provides insights into origin of weedy rice and rice domestication[J].Journal of Heredity,2014,105(2): 276-287.
[24] SUNDSTR?魻M J F,VACULOVA A,SMERTENKO A P,et al.Tudor staphylococcal nuclease is an evolutionarily conserved component of the programmed cell death degradome[J].Nature Cell Biology,2009,11(11):1347-1354.
[25] CHEN X. A microRNA as a translational repressor of APETALA2 in Arabidopsis flower development[J].Science,2004, 303(5666):2022-2025.
[26] TAKAGI H,TAMIRU M,ABE A,et al. MutMap accelerates breeding of a salt-tolerant rice cultivar[J].Nature Biotechnology,2015,33(5):445-449.
