高温胁迫对圆齿野鸦椿幼苗生理代谢的影响
2017-07-06刘福辉
刘福辉
(福建省林业调查规划院,福建 福州 350003)
植物在生长发育过程中会受到各种非生物因子的胁迫,其中温度是影响植物生长的主要因子之一。高温胁迫下植物细胞内部酶促反应会发生剧烈变化,酶活性变化会引发系列植物细胞代谢失衡,使光合作用和呼吸过程受到影响[1]。植物通过调节新陈代谢进行热胁迫响应,如产生能促进蛋白和细胞重建的物质、调节渗透来保持细胞膨胀、改变体内氧化系统来重建细胞平衡和稳态等。当高温胁迫不断加强,将导致植物细胞膜损伤、光合作用受到抑制、细胞老化死亡,甚至植物表现出外观形态的热害症状[2]。圆齿野鸦椿(Euscapiskonishii)为省沽油科(Staphyleaceae)野鸦椿属(Euscapis)的常绿小乔木,分布福建、广东、广西、江西等省区[3]。其树形婆娑优美,春夏之际,新枝鲜红,老枝暗红,黄绿花序多而密;秋季,蓇葖果由绿转红,红果期跨越秋、冬和翌年春三季。成熟果实沿内腹缝线开裂,鲜红色内果皮外翻呈两瓣蝴蝶翅状,观果期长达8个月,是一极具潜力的园林绿化树种[4,5]。作为传统民间药用植物,其根、果性温,味微苦,有祛风除湿,止血止痛,发表散寒等功效[6]。目前,对圆齿野鸦椿的研究集中于生物学特性、种苗繁育技术、生理生化特性以及开发利用等方面[5,7~8],未见有关高温胁迫下圆齿野鸦椿幼苗的光合特性、抗氧化酶和碳水化合物代谢方面的报道。
1 材料与方法
1.1 植物材料
2016年3月,将经过1年沙藏处理的裂口5%左右的圆齿野鸦椿种子播种于上口径15 cm、下口径10 cm、高10 cm的塑料花盆中,栽培基质为炭化谷壳、黄心土和泥炭土,按体积比2∶1∶1混合均匀,每盆均匀撒播30~40粒种子。放入福建农林大学田间试验温室中育苗。
1.2 试验设计与处理
每盆留苗25~30株,待苗高约15 cm时,选健康、无病虫害、长势好的作为试验苗,共计20盆。转入人工气候箱进行培养。生长条件为昼夜温度25℃/20℃,相对湿度65%~75%,光周期为10h.d-1。预培养7d后进行温度处理,其中一个生长箱昼夜温度为25℃/20℃(对照),另一个生长箱昼夜温度为35℃/25℃(高温处理),每个处理10次重复,相对湿度仍为65%~75%,光周期为10h.d-1。各处理盆栽土壤含水量保持30%~35%。
1.3 测定指标与方法
1.3.1 叶绿素含量测定
温度处理后,分别于第1天、第2天、第3天和第4天,取正常生长的新鲜幼苗叶片各0.2 g,剪碎后研磨成匀浆,彻底离心后选用分光光度法对类胡萝卜素的含量加以测定,而叶绿素a、叶绿素b、总叶绿素含量的测定则采用Arnon的方法[9]。
1.3.2 叶绿素荧光参数测定
分别于高温处理20 min、25 min、30 min、35 min后,随机选取各组幼苗10片叶,采用FMS-2型荧光仪进行测定。叶绿素荧光测定过程中,提前将试验材料暗处理30 min,之后再利用12 000 μmol·m-2·s-1的饱和脉冲光照射,试验完毕后测定PSII最大的光化学量子产量Fv/Fm、光化学猝灭系数qP、PSII系数和1- qP[9]。
1.3.3 碳水化合物的提取
待培养1个月后,取若干叶片样本,各试验组中摘取0.1 g叶片研磨后置于试管中,15 min后向其中倒入8 ml浓度为75%的乙醇进行80℃水浴30 min,后全部离心再反复进行提纯,用活性炭对提取液中的色素进行吸附。将试管烘干,加入3 ml蒸馏水后摇匀放置于沸水中加热片刻,待彻底冷却后向其中滴入3 ml浓度9 mol·L-1高氯酸反复进行提取后将上清液取出待用[9]。
1.3.4 酶液的提取
待培养1个月后,各试验组中分别摘取0.2 g新鲜叶片,研磨后向其中注入2 mL 25 mM的HEPES和2.5%的非溶性PVP,研磨后离心15 min取其上清液待检测酶液[9]。
1.4 数据处理
采用EXCELL 2003和SPSS.19软件进行数据分析。
2 结果与分析
2.1 对碳水化合物代谢的影响
高温胁迫下圆齿野鸦椿叶片中蔗糖、可溶性糖、还原性糖比对照组分别提高47.8%、9.8%、18.6%;但淀粉含量仅为非高温胁迫下淀粉含量的68.5%。淀粉含量的下降可能是由于温度升高导致圆齿野鸦椿幼苗叶片内淀粉发生降解,直接导致了蔗糖、还原性糖等可溶性糖含量直接升高,其作用在于使植物细胞膜的成分更加固定,一定程度上能够降低高温胁迫对圆齿野鸦椿幼苗的危害。碳水化合物含量的变化能一定程度上体现当前时间段圆齿野鸦椿幼苗光合作用效率的高低。一般情况下,光合作用效率的提升能够使得植物体内碳水化合物含量得到提高,而本试验中碳水化合物含量的变化也证明了试验过程中圆齿野鸦椿幼苗的光合作用效率发生了变化。
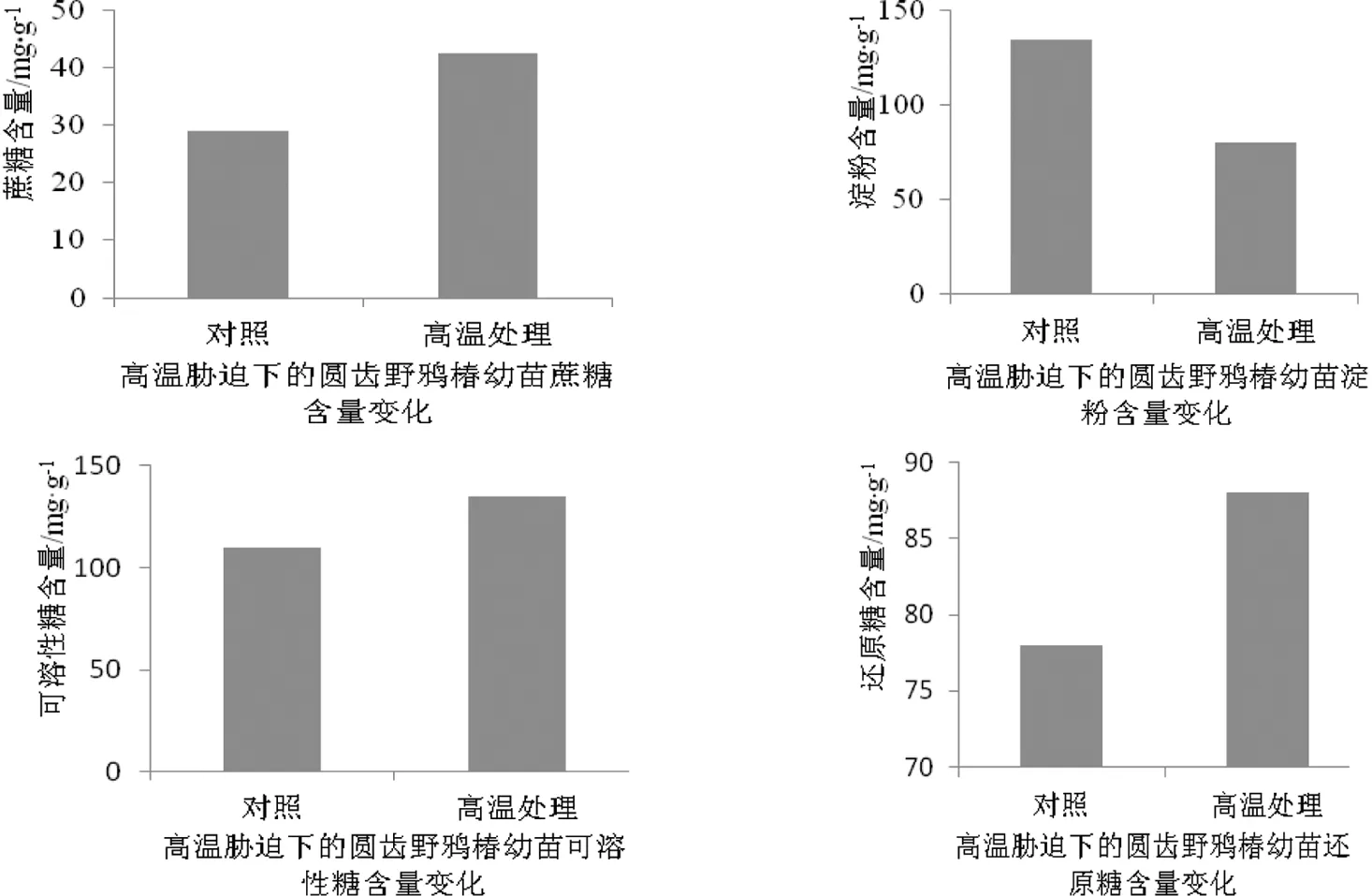
图1 高温胁迫对圆齿野鸦椿碳水化合物含量的影响
2.2 对叶绿素含量的影响
高温胁迫下,圆齿野鸦椿幼苗叶片内叶绿素a含量、叶绿素b含量、总叶绿素含量和类胡萝卜素含量比对照组降低了20.5~31.5%、20~30%、22.5~30.5%和18.5~29.5%,降幅依次为26%、25%、26.5%和24%。随着高温胁迫的不断持续,圆齿野鸦椿幼苗体内叶绿素和类胡萝卜素的含量均呈现先降后升的趋势,但总体是明显下降。
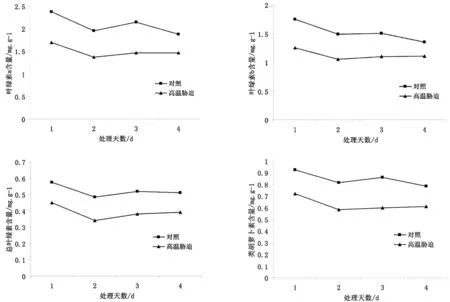
图2 高温胁迫对圆齿野鸦椿幼苗叶绿素、类胡萝卜素含量的影响
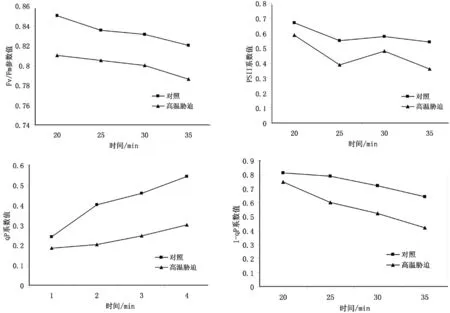
图3 高温胁迫对圆齿野鸦椿叶片叶绿素荧光参数的影响
2.3 对叶绿素荧光参数的影响
Fv/Fm代表PSII最大的光化学量子产量,qP代表光化学猝灭系数,反映PSII中心电化学反应的能力强弱,系数越大,则表明电子传递活性的强度也就越高。高温胁迫导致了Fv/Fm呈下降趋势,PSII系数也呈下降趋势,但是在25~30 min的过程中对照组和高温胁迫之后的圆齿野鸦椿都有些许波动,表明高温胁迫下,圆齿野鸦椿幼苗同化力(NADPH、ATP)的形成受阻,影响了植物对碳的固定和同化。随着胁迫时间的延长,qP逐渐降低,而1-qP逐渐升高,表明PSII的还原状态在高温胁迫下相对较高。
2.4 对抗氧化酶活性的影响
抗氧化酶是动植物体内抵抗细胞衰老的重要物质,其机理在于组织细胞与氧气的直接接触,从而减缓细胞氧化速度。高温胁迫下圆齿野鸦椿叶片、根茎中超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)的活性均大于对照组;叶片中愈创木酚过氧化物酶(GPX)的活性大于对照组,但根茎中GPX的活性比对照组小。
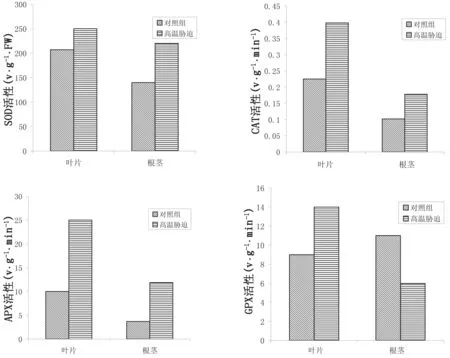
图4 高温胁迫对圆齿野鸦椿叶片和根茎抗氧化酶活性的影响
3 讨论
3.1 高温对圆齿野鸦椿幼苗碳水化合物代谢的影响
高温胁迫可引起植物细胞膜渗透性增大而导致电解质渗透。为防止电解质过多外渗,植物通过调节物质的种类和含量以适应逆境,其中可溶性糖是植物组织中的渗透调节物质之一[10]。本研究结果表明,高温胁迫下,圆齿野鸦椿幼苗中可溶性糖含量比对照组高。当可溶性糖含量较高时,可以降低细胞的渗透势从而维持细胞的膨压,防止细胞内大量的被动脱水。淀粉是植物主要的能量储藏物质,是在叶绿体或淀粉体中通过一系列酶促反应合成的,在高温胁迫下,植物呼吸作用加强从而加快淀粉分解[11]。本研究结果中,高温胁迫下淀粉的含量明显比对照组低,说明淀粉在圆齿野鸦椿幼苗的耐热性中起了重要作用。蔗糖是植物糖分的重要组成部分,它是植物光合作用的主要产物,是碳运输的主要形式,也是细胞代谢的调控因子[12]。高温胁迫下,蔗糖含量高于对照组,可能是因为幼苗为了适应逆境,加快光合作用。
一般而言,温度对于植物生长具有十分重要的影响,过高或过低都无法满足植物生长。圆齿野鸦椿幼苗生长过程中,其新陈代谢的发生与体内一系列的酶促反应息息相关,当温度未达到抑制酶促反应发生所需的最高温度,则当温度升高时,幼苗体内的酶活性提升,所引发的酶促反应相对更加剧烈。在此情况下,圆齿野鸦椿幼苗叶片内蔗糖、还原性糖、可溶性糖含量分别提升,而随着温度的不断升高,淀粉开始分解,进而提升蔗糖、还原性糖、可溶性糖的整体含量,而在此过程中淀粉含量逐渐降低。
3.2 高温对圆齿野鸦椿幼苗叶片光合作用的影响
光合作用是一切生物直接或间接的能量来源,是植物进行物质转换和能量代谢的关键。叶绿素荧光技术通过对植物光合过程中荧光特性的探测可了解植物的生长、病害及受胁迫等生理状态[13]。研究表明正常生长植物叶片中光能转化效率达到0.85。当对正常生长的植物进行高温胁迫时,光能转化效率会降低,反映在高温胁迫过程中植物叶片细胞的光合作用受到抑制[14]。PSII指标反映光合作用过程中线性电子传递电子流的能力强弱,在正常的光合作用过程中线性电子传递电子流的能量对于植物的正常生长发育至关重要。本研究中,在高温胁迫下,PSII、Fv/Fm和qP明显降低,而1-qP增大,这与Abdul K等人的研究结果一致[15]。表明光合电子传递以分子态氧为受体的支路反应增强和PSII的部分失活,抑制光合碳代谢电子供应,导致光合作用受阻[16],同时叶片细胞的有机物合成能力也受到抑制。
叶绿素作为植物进行光合作用的主要色素,其含量在一定程度上反映了叶片光合作用能力的大小[17]。本研究表明,高温胁迫导致圆齿野鸦椿幼苗叶片叶绿素和类胡萝卜素含量明显降低,原因可能有3点:一是高温胁迫降低叶绿素合成速率。大多数植物即使在短时间(15~60 min)的高温(35~40℃)胁迫下,其光合作用就会受到明显的抑制[18]。二是高温胁迫导致活性氧的积累,加速叶绿素的降解[19]。三是本研究中叶绿素a、b下降的不平衡性,导致叶片光合作用的降低,这与张桂莲[20]等人的研究结果相同。类胡萝卜素既是光合色素,又是细胞内源抗氧化剂,一方面吸收光能并传递给反应中心,另一方面可以吸收剩余能量,淬灭活性氧,防止膜质过氧化,因此类胡萝卜素的减少,也可导致叶绿素含量的降低[21]。
3.3 高温对圆齿野鸦椿幼苗抗氧化酶活性的影响
植物正常生长的细胞内自由基的产生与清除处于动态平衡状态,自由基水平很低,不会伤害细胞。当植物受到高温胁迫时,植物体内的活性氧含量增加,若不及时清除,必导致氧化胁迫,膜系统受到破坏,打破自由基的平衡,导致植物体内的自由基清除机制遭到破坏[22]。为避免这种伤害,植物在长期的进化过程中,形成一套活性氧的清除体系,主要包括酶促防御系统和非酶促防御系统[23]。酶促防御系统主要由SOD、AXP、GPX和CAT构成,其中,CAT通过催化细胞内过氧化氢的分解,防止膜质过氧化[24];APX利用抗坏血酸为电子供体的H2O2清除剂,大量研究表明叶绿体中的H2O2由APX清除[25];GPX是动物细胞中的一种重要H2O2清除剂,但是现在的研究发现在植物中GPX也具有保护细胞免受氧化伤害的作用[26]。在本研究中,高温胁迫下,圆齿野鸦椿叶片中SOD、AXP、GPX和CAT的活性提高,表明叶片对高温胁迫产生的适应反应;根茎SOD、AXP和CAT的活性也提高了,但GPX活性却呈现下降趋势,可能是根茎还不适应高温环境。
[1] 王利琳, 庞基良, 胡江琴, 等.温度对植物成花的影响 [J].植物学通报, 2002, 19 (2): 5-8.
[2] 屠小菊, 汪启明, 饶力群. 高温胁迫对植物生理生化的影响 [J]. 湖南农业科学, 2013 (7): 28-30.
[3] 邹小兴, 刘宇, 邹双全. 圆齿野鸦椿叶及枝化学成分初步研究 [J]. 中国野生植物资源, 2016, 35(1): 70-72.
[4] 敖小朋, 徐耀东, 童志刚.野生赏药兼用植物野鸦椿 [J].广西农学报, 2009, 24 (1): 45-46.
[5] 孙维红,袁雪艳,吴玲娇,等.圆齿野鸦椿开花动态特征和繁育系统 [J]. 植物生理学报,2017,53 (12): 2215-2221.
[6] 覃嘉佳, 龙云英.圆齿野鸦椿扦插繁殖技术 [J].林业科技开发, 2007, 21 (3): 71-73.
[7] 何碧珠, 何官榕, 邹双全.圆齿野鸦椿叶片的植株再生及快速繁殖 [J].福建农林大学学报(自然科学版), 2010, 39 (3): 257-262.
[8] 黄铭星, 邹双全, 陈琳, 等.施用人工菌剂对圆齿野鸦椿幼苗移栽生长的影响 [J].福建林学院学报, 2013, 33 (1): 25-27.
[9] 李合生.植物生理生化试验原理和技术 [M].北京: 高等教育出版社, 2000.
[10] Patton A J, Cunningham S M, Volenec J J, et al. Differences in Freeze Tolerance of Zoysiagrasses: II. Carbohydrate and Proline Accumulation All rights reserved. No part of this periodical may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or any information storage and retrieval system, without permission in writing from the publisher. Permission for printing and for reprinting the material contained herein has been obtained by the publisher [J]. Crop Science, 2007, 47(5): 2170-2181.
[11] 王日明, 熊兴耀. 高温胁迫对黑麦草生长及生理代谢的影响 [J]. 草业学报, 2016, 25(8): 81-90.
[12] 张建波, 金云峰, 王莎莎, 等. 生长温度对不同生育期烟草蔗糖代谢的影响 [J]. 基因组学与应用生物学, 2015 (10): 2225-2244.
[13] 冯伟, 李晓, 王永华, 等. 小麦叶绿素荧光参数叶位差异及其与植株氮含量的关系 [J]. 作物学报, 2012, 38(4): 657-664.
[14] 贾虎森, 李德全, 韩亚琴.高等植物光合作用的光抑制研究进展 [J].植物学通报, 2000, 17 (3): 218-224.
[15] Karim A, Fukamachi H, Hidaka T. Photosynthetic performance of Vigna radiata L. leaves developed at different temperature and irradiance levels [J]. Plant Science, 2003, 164(4): 451-458.
[16] 彭长连, 林植芳. 光氧化胁迫下几种植物叶片的超氧自由基产生速率和光合特性 [J]. 植物生理学报, 2000, 26(2): 81-87.
[17] 朱先灿, 宋凤斌, 徐洪文. 低温胁迫下丛枝菌根真菌对玉米光合特性的影响 [J]. 应用生态学报, 2010, 21(2): 470-475.
[18] Xu Q, Huang B. Lowering soil temperatures improves creeping bentgrass growth under heat stress[J]. Crop Science, 2001, 41(6): 1878-1883.
[19] 徐娜婷, 刘玉民, 刘亚敏, 等. 引种花卉紫串扦插苗对高温胁迫的生理响应 [J]. 草业学报, 2012, 21(5): 257-263.
[20] 张桂莲, 陈立云, 张顺堂, 等. 抽穗开花期高温对水稻剑叶理化特性的影响 [J].中国农业科学, 2007(7):1345-1352.
[21] 汪炳良. 高温胁迫对早熟花椰菜叶片抗氧化系统和叶绿素及其荧光参数的影响 [J]. 中国农业科学, 2004, 37(08): 1245-1245.
[22] 张悦, 井维霞, 刘修堂, 等. 不同浓度安打对烟草幼苗 MDA, CAT, APX 和 GPX 的影响 [J]. 生物灾害科学, 2013 (1): 22-25.
[23] Inzé D, Van Montagu M. Oxidative stress in plants [J]. Current Opinion in Biotechnology, 1995, 6(2): 153-158.
[24] Van Breusegem F, Vranová E, Dat J F, et al. The role of active oxygen species in plant signal transduction [J]. Plant Science, 2001, 161(3): 405-414.
[25] 孙小妹, 张涛, 陈年来, 等. 土壤水分和氮素对春小麦叶片抗氧化系统的影响 [J].干旱区研究, 2011(2):205-214.
[26] 王艳, 李建龙, 邓蕾, 等. H2O2 对沟叶结缕草抗氧化系统及其耐寒性的影响 [J]. 中国草地学报, 2010, 32(2): 92-97.
