山乌桕对淹水胁迫的生理响应
2017-06-21陈敏旗王小德
陈敏旗,王小德
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
山乌桕对淹水胁迫的生理响应
陈敏旗,王小德*
(浙江农林大学 风景园林与建筑学院,浙江 临安 311300)
为研究淹水胁迫对山乌桕(Sapiumdiscolor)生理特性的影响,该文以乌桕(Sapiumsebiferum)为对照,通过人工模拟淹水环境,对山乌桕和乌桕的叶绿素含量、相对电导率、超氧化物歧化酶(SOD)和过氧化物歧化酶(POD)活性、可溶性糖和可溶性蛋白含量等进行比较研究。结果表明:随着处理时间的延长,山乌桕、乌桕的叶绿素含量下降;两者细胞膜透性均受到不同程度的损伤,电导率上升;SOD活性增强,POD活性下降,其中,山乌桕的SOD、POD活性均低于乌桕;两者可溶性糖、可溶性蛋白、脯氨酸含量均呈上升趋势,但乌桕的上升幅度显著高于山乌桕。表明山乌桕有一定的耐涝性,但其耐涝性不如乌桕。
山乌桕; 淹水胁迫; 叶绿素含量; 渗透调节物质
山乌桕为大戟科乌桕属落叶乔木或灌木[1],具有树干直立、病虫害较少、季相变化丰富等特点,已成为近年来浙江省风景林推广应用的优良珍贵乡土树种之一。目前,国内外关于乌桕的耐涝性研究较为广泛,而对山乌桕的研究主要集中在酿蜜、引种驯化、推广应用、生物分布及群落特征、育苗造林等方面,对其耐涝性报道较少。本试验通过人工模拟淹水环境,以乌桕为对照,研究山乌桕耐涝生理变化过程,为山乌桕在园林中的应用和风景林的营造上寻找更为广阔的立地条件提供理论依据。
1 材料与方法
1.1 试验材料
试验在浙江农林大学植物生理实验室进行。试验材料为2年生山乌桕实生苗(来自浙江龙泉省级种苗基地)、2年生乌桕实生苗(来自浙江临安天目山林场种苗基地)。于2016年2月初,选择生长相对一致的苗木进行盆栽(盆口径21 cm,高20 cm),盆土按原土∶基质体积比1∶1配置,置于浙江农林大学园林学院植物实习基地内,生长期间进行正常养护管理。
1.2 处理设计
试验开始于7月5日,挑选长势相对一致的山乌桕和乌桕进行淹水处理,将山乌桕和乌桕放入规格相同的塑料水桶内,土壤含水量完全饱和,试验过程中不断加水,以保持淹水深度超过盆土面,模拟自然条件下的涝害环境。每个处理重复3次,7 d为1个处理周期,检测山乌桕、乌桕生理指标的变化。
1.3 指标测定
叶绿素含量的测定:选取用于测定光和色素的叶片,采用浸提法[2]测定;叶片相对电导率测定采用抽气法[2];SOD活性测定采取氮蓝四唑(NBT)法[3];POD活性参照愈创木酚比色法[3];可溶性糖质量分数测定采用蒽酮比色法[3];可溶性蛋白含量测定采用考马斯亮蓝染色法[3];脯氨酸含量测定参照酸性茚三酮法[3]。
1.4 数据处理
采用WPS和SPSS 20进行数据统计与分析,用OriginPro 7.5绘图。
2 结果与分析
2.1 淹水胁迫对山乌桕、乌桕叶绿素含量的影响
由图1可知,淹水处理下山乌桕、乌桕叶片的叶绿素含量随着胁迫时间延长而下降,其中乌桕下降速率总体比山乌桕小,乌桕叶绿素含量总体高于山乌桕。第0天,乌桕叶绿素含量比山乌桕叶绿素含量低2.2%,第7、14、21、28天,乌桕叶片叶绿素含量比山乌桕叶片叶绿素含量分别高5.6%、7.4%、15.1%、23.2%;山乌桕、乌桕第28天叶绿素含量比第0天分别低49.2%、32.3%。
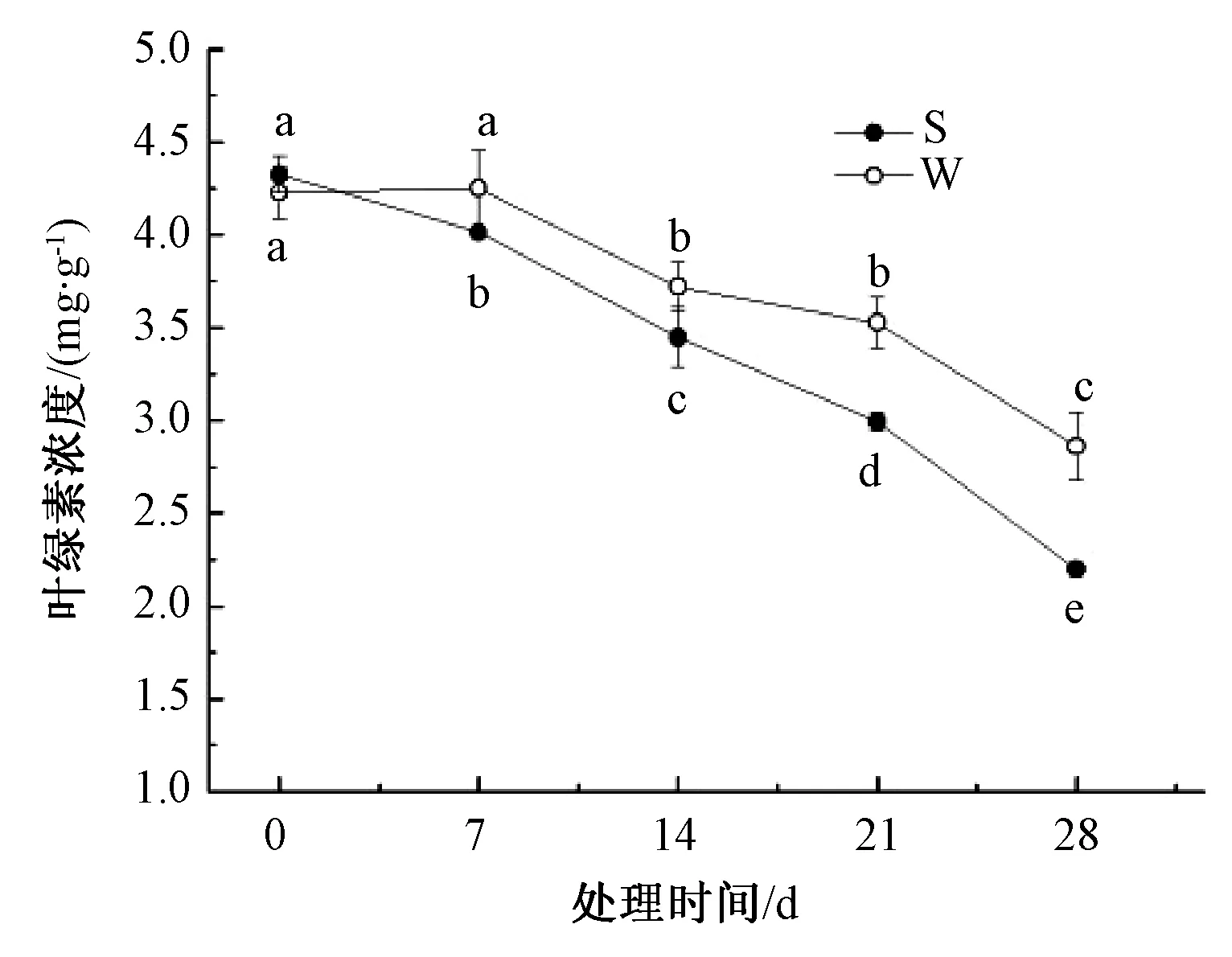
S代表山乌桕,W代表乌桕。同一时间不同处理无相同小写字母表示差异显著(P<0.05)。图2~7同图1 淹水胁迫对山乌桕、乌桕叶绿素含量的影响
2.2 淹水胁迫对山乌桕、乌桕叶片相对电导率的影响
山乌桕、乌桕叶片相对电导率在淹水胁迫下整体呈现上升的趋势,说明细胞膜受到一定程度的伤害。由图2可知,7~14 d,乌桕叶片相对电导率呈下降趋势,可能是乌桕自身调节机制对淹水胁迫有一定的抗性。山乌桕、乌桕叶片相对电导率上升幅度不大,整个处理期间分别上升了35.8%和35.9%。
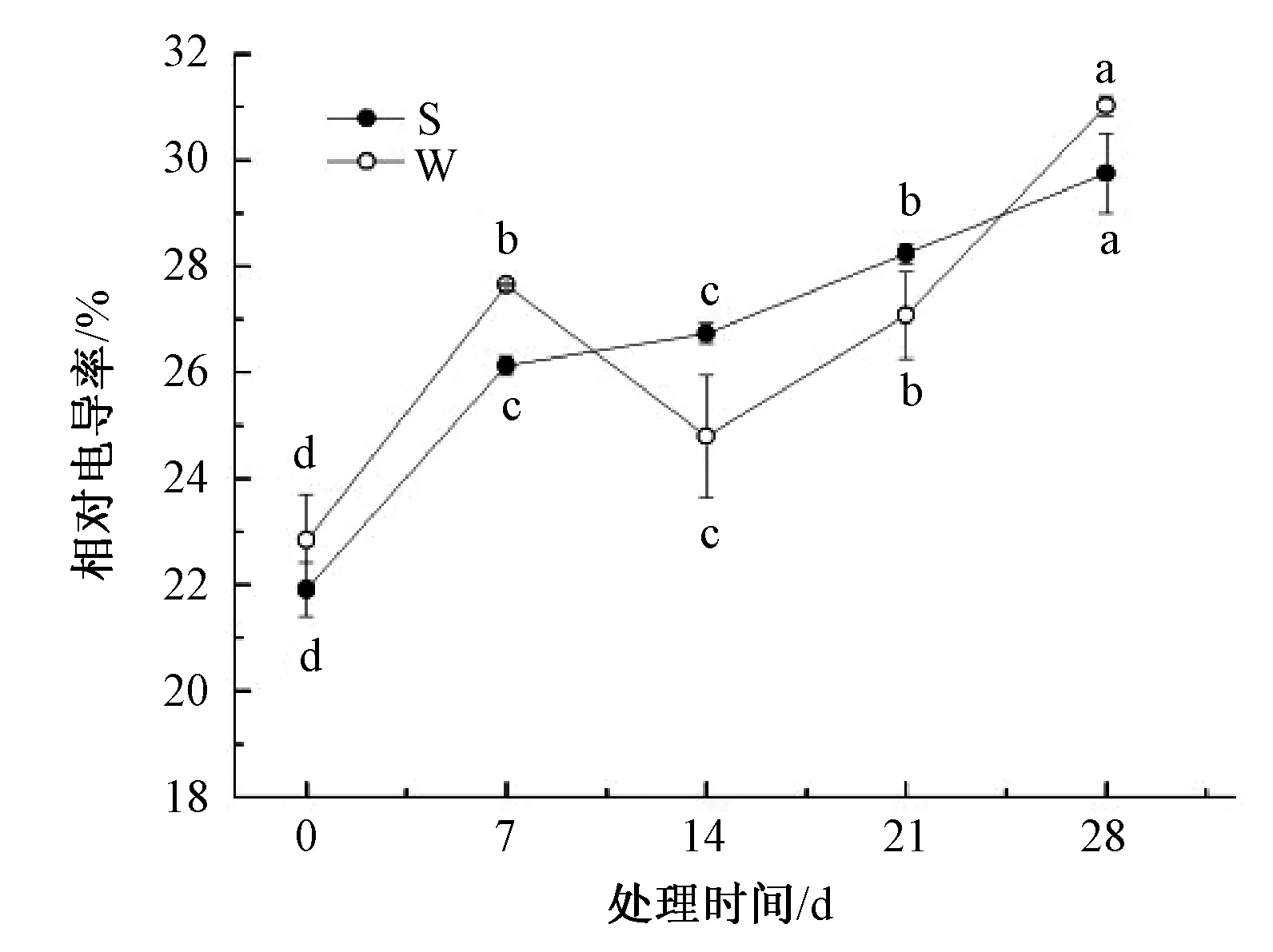
图2 淹水胁迫对山乌桕、乌桕相对电导率的影响
2.3 淹水胁迫对山乌桕、乌桕叶片SOD活性的影响
由图3可知,淹水胁迫下,山乌桕、乌桕叶片SOD活性整体都呈逐渐上升的趋势,胁迫前山乌桕叶片SOD活性比乌桕高16.1%,胁迫前中期山乌桕SOD活性一直高于乌桕,21~28 d山乌桕SOD活性迅速下降,乌桕SOD活性则持续上升。
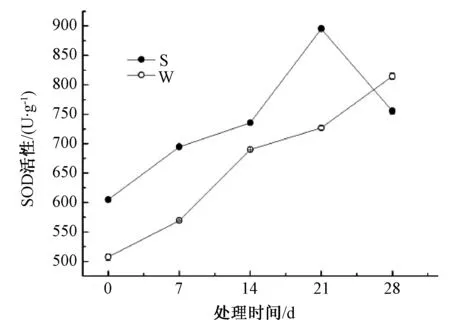
图3 淹水胁迫对山乌桕、乌桕SOD活性的影响
2.4 淹水胁迫对山乌桕、乌桕叶片POD活性的影响
由图4可知,淹水胁迫下,山乌桕、乌桕POD活性变化总体呈下降的趋势,整个处理周期中分别下降了30.8%、80.3%。胁迫初期,乌桕POD活性比山乌桕高48.9%,0~7 d,山乌桕POD活性上升了4.2%,乌桕POD活性则下降了14.3%;7~14 d,山乌桕和乌桕POD活性都迅速下降,分别比7 d下降了47.0%、54.3%;14~28 d,山乌桕POD活性变化趋势和乌桕基本一致。
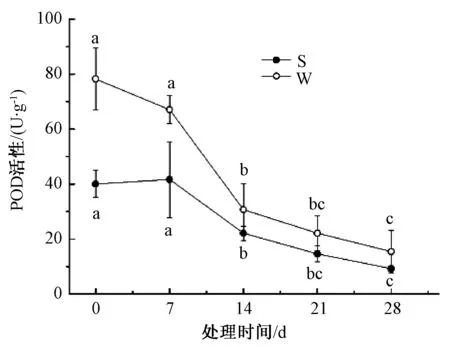
图4 淹水胁迫对山乌桕、乌桕POD活性的影响
2.5 淹水胁迫对山乌桕、乌桕叶片可溶性糖含量的影响
由图5可知,淹水胁迫前,山乌桕、乌桕可溶性糖含量基本相同。0~7 d,山乌桕、乌桕可溶性糖含量都呈上升趋势,分别上升了35.7%、75.3%,且第7 天乌桕可溶性糖含量显著高于山乌桕;7~14 d,山乌桕可溶性糖含量上升,而乌桕则小幅下降;14~21 d,山乌桕、乌桕可溶性糖含量都有所上升,山乌桕上升速率比乌桕快;胁迫末期乌桕可溶性糖含量持续上升,28 d比21 d上升了10.1%,山乌桕可溶性糖含量则逐渐下降,28 d比21 d下降了23.1%。
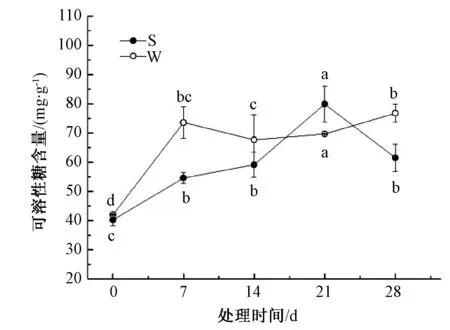
图5 淹水胁迫对山乌桕、乌桕可溶性糖含量的影响
2.6 淹水胁迫对山乌桕、乌桕叶片可溶性蛋白含量的影响
由图6可知,淹水胁迫下,山乌桕可溶性蛋白变化较为复杂,在一定范围内呈波动变化;乌桕可溶性蛋白含量总体呈上升趋势。乌桕可溶性蛋白含量在整个处理周期内都高于山乌桕,28 d时,乌桕可溶性蛋白含量是山乌桕的3.0倍。0~7 d,山乌桕、乌桕可溶性蛋白含量都呈上升趋势,7~14 d,山乌桕可溶性蛋白含量下降了38.5%,而乌桕可溶性蛋白则上升了54.1%;14~21 d,山乌桕可溶性蛋白含量快速上升,乌桕可溶性蛋白含量则小幅上升;21~28 d,乌桕可溶性蛋白含量持续缓慢上升,山乌桕可溶性蛋白快速下降。
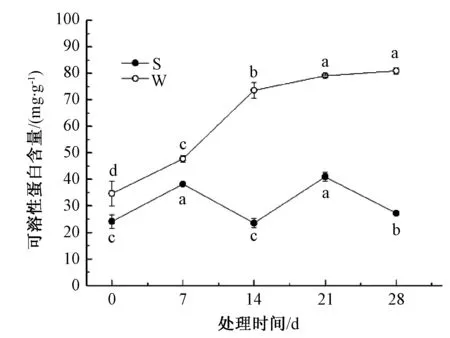
图6 淹水胁迫对山乌桕、乌桕可溶性蛋白含量的影响
2.7 淹水胁迫对山乌桕、乌桕叶片脯氨酸含量的影响
由图7可知,淹水处理下山乌桕、乌桕脯氨酸整体呈上升趋势,28 d分别比0 d高75.9%、79.2%。胁迫处理第7天,山乌桕脯氨酸含量显著高于乌桕,是乌桕脯氨酸含量的3.2倍;7~14 d,山乌桕脯氨酸小幅下降,乌桕脯氨酸快速上升,到第14天时,两者的脯氨酸含量持平;14~21 d,两者上升速率相差不大,到第21天时,二者脯氨酸含量均在100 mg·g-1左右;21~28 d,山乌桕脯氨酸含量有所下降,乌桕脯氨酸含量则持续上升。
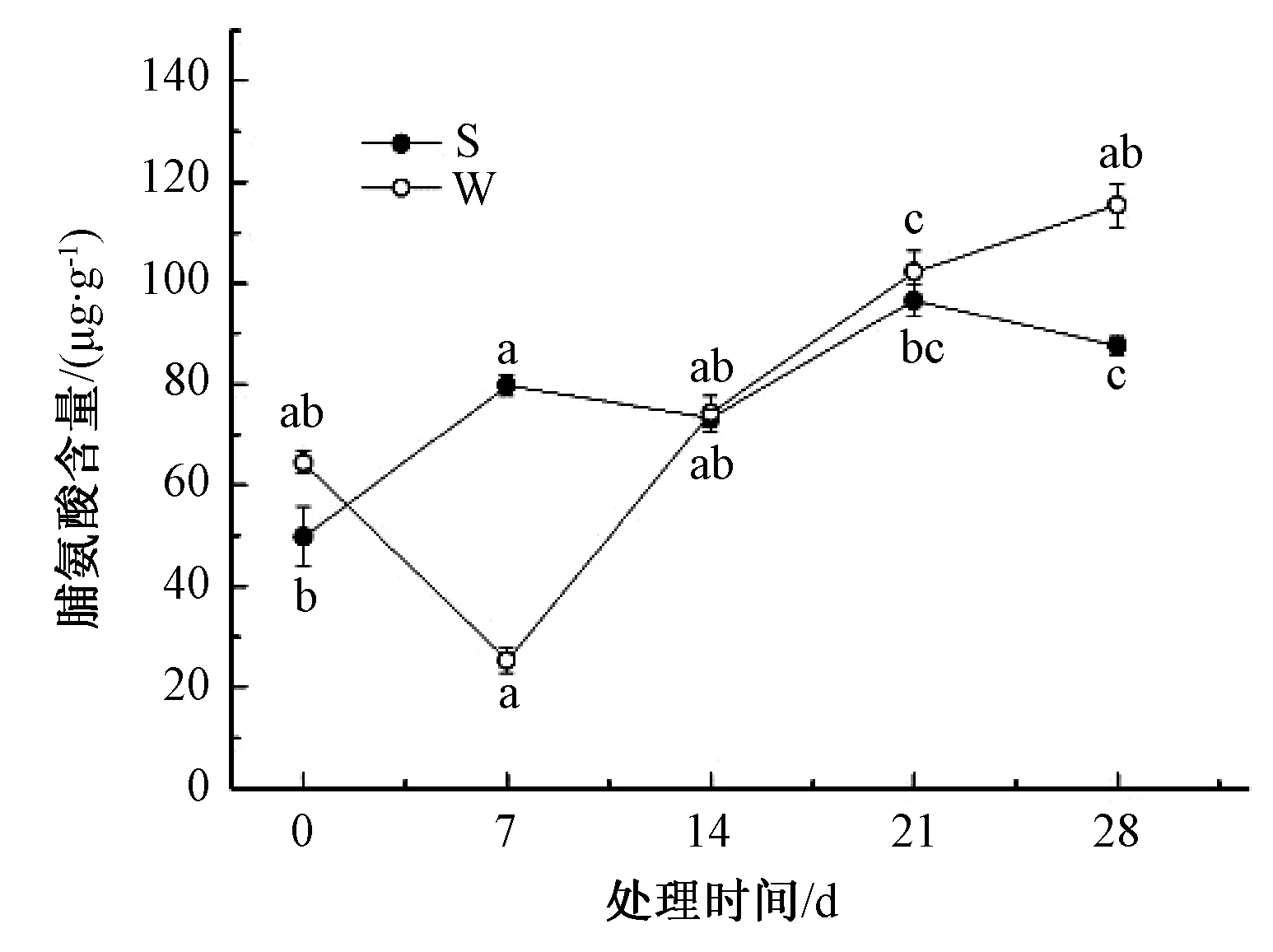
图7 淹水胁迫对山乌桕、乌桕叶片脯氨酸含量的影响
3 小结与讨论
随着淹水时间的延长,气体扩散受限,叶片细胞膜脂过氧化加剧,叶绿素降解,叶片相对电导率上升,体内保护酶系统平衡被打破。为了适应淹水环境,植物通过生理生化机制的调节来保证淹水条件下的生命活动,如细胞通过调节渗透物质的含量来保持渗透势的平衡;细胞内各种抗氧化酶活性增加,以清除氧自由基,避免或者减轻细胞受到伤害。因此,植物叶片渗透调节物质含量及保护酶活性的高低在一定程度上反映了植物的耐涝性强弱[4-5]。本试验表明,淹水胁迫时,山乌桕和乌桕的叶绿素含量下降、相对电导率持续上升,说明两者均受到一定程度的伤害,这与王晓荣等[6]和曹福亮等[7]在乌桕研究中的结果一致,尹冬梅等[8]在菊花及何斌源等[9]在红树植物研究中在也有相似结论。但胁迫中期山乌桕相对电导率高于乌桕,说明山乌桕叶片受到的伤害可能大于乌桕叶片。
淹水胁迫时,植物体内会产生并积累有毒物质氧自由基,植物会启动抗氧化系统来清除氧自由基[10]。山乌桕和乌桕受到淹水胁迫时,SOD活性升高,可能是胁迫导致植物体内产生活性氧的积累诱导SOD活性提高,从而减少活性氧带来的破坏。胁迫后期,山乌桕SOD活性有所下降,可能是因为活性氧的积累超过了抗氧化系统的清除能力范围。两者的POD活性与SOD活性变化呈相反趋势,胁迫14 d时,POD活性保持在较低水平,可能是长期胁迫下山乌桕受到伤害的主要原因之一。这与王哲宇等[11]对朴树的研究结果和汪贵斌等[12]对喜树的研究结果相似。在整个胁迫过程中,乌桕叶片的SOD和POD活性一直高于山乌桕,可能是乌桕的耐涝性强于山乌桕。
可溶性蛋白、可溶性糖、脯氨酸含量都是植物细胞重要的渗透调节物质。在胁迫条件下,植物体内蛋白质含量的提高是植物适应逆境胁迫的一种表现,可作为植物相对抗性的一种指标[13]。糖酵解是植物对缺氧胁迫适应的基本功能,维持糖酵解途径需要源源不断的可溶性糖的供应[14]。所以,淹水处理下植物可溶性糖含量是指示植物耐涝性的重要指标。本试验发现,山乌桕叶片可溶性蛋白、可溶性糖、脯氨酸含量均随着处理时间的延长呈增长趋势,这些物质能调节体内代谢,因而能在一定程度上减轻淹水的危害。这与连洪燕[15]对石楠属植物的研究结果及张晓平等[16]对鹅掌楸属植物的研究结果一致。几乎整个处理过程中乌桕的渗透调节物质含量都高于山乌桕,在一定程度上说明乌桕的耐涝性强于山乌桕。
[1] 马金双. 中国植物志[M]. 北京: 科学出版社, 1997.
[2] 王学奎. 植物生理生化实验原理与技术[M]. 北京: 高等教育出版社, 2006.
[3] 李合生. 植物生理生化实验[M].北京: 高等教育出版社, 2000.
[4] 曾德静, 王铖, 刘军, 等. 水涝胁迫下海州常山形态和生理指标的变化[J]. 浙江农林大学学报, 2013, 30(2): 172-178.
[5] YIN D M, CHEN S M, CHEN F, et al. Morpho-anatomical and physiological responses of twoDendranthemaspecies to waterlogging [J]. Environmental and Experimental Botany, 2010, 68(2): 122-130.
[6] 王晓荣, 胡兴宜, 唐万鹏, 等. 模拟长江滩地水淹胁迫对3种树种幼苗生理生态特征的影响[J]. 东北林业大学学报, 2015 (1): 45-49.
[7] 曹福亮, 蔡金峰, 汪贵斌, 等. 淹水胁迫对乌桕生长及光合作用的影响[J]. 林业科学, 2010, 46(10): 57-61.
[8] 尹冬梅, 陈发棣, 陈素梅. 涝渍胁迫下5种菊花近缘种属植物生理特性[J]. 生态学报, 2009, 29(4): 2143-2148.
[9] 何斌源, 赖廷和, 陈剑锋, 等. 两种红树植物白骨壤(Avicenniamarina)和桐花树(Aegicerascorniculatum)的耐淹性[J]. 生态学报, 2007, 27(3): 1130-1138.
[10] 汪贵斌, 曹福亮, 张晓燕, 等. 涝渍胁迫对不同树种生长和能量代谢酶活性的影响[J]. 应用生态学报, 2010, 21(3): 590-596.
[11] 王哲宇, 童丽丽, 汤庚国. 淹水胁迫对朴树幼苗形态及生理特性的影响[J]. 林业科技开发, 2013, 27(4): 99-103.
[12] 汪贵斌, 蔡金峰, 何肖华. 涝渍胁迫对喜树幼苗形态和生理的影响[J]. 植物生态学报, 2009, 33(1): 134-140.
[13] 潘澜, 薛立. 植物淹水胁迫的生理学机制研究进展[J]. 生态学杂志, 2012, 31(10): 2662-2672.
[14] 谭淑端, 朱明勇, 张克荣, 等. 植物对水淹胁迫的响应与适应[J]. 生态学杂志, 2009, 28(9): 1871-1877.
[15] 连洪燕. 淹水胁迫对三种石楠属植物幼苗可溶性糖和脯氨酸含量的影响[J]. 北方园艺, 2012 (20): 63-66.
[16] 张晓平, 方炎明, 陈永江. 淹涝胁迫对鹅掌楸属植物叶片部分生理指标的影响[J]. 植物资源与环境学报, 2006, 15(1): 41-44.
(责任编辑:侯春晓)
2017-03-09
陈敏旗(1992—),女,江西上饶人,硕士研究生,研究方向为植物景观设计与评价、植物抗逆性生理,E-mail: 1115122576@qq.com。
王小德(1965—),男,浙江临安人,教授,博士,研究方向为园林植物引种与应用、植物造景和生态园林等,E-mail: wxd65@zafu.edu。
10.16178/j.issn.0528-9017.20170533
S687.1
A
0528-9017(2017)05-0829-04
文献著录格式:陈敏旗,王小德. 山乌桕对淹水胁迫的生理响应[J].浙江农业科学,2017,58(5):829-832.
