猪可代谢营养评定新体系研究进展
2017-05-12赵胜军武汉轻工大学动物营养与饲料科学湖北省重点实验室武汉430023动物营养与饲料安全湖北省协同创新中心武汉430023
赵胜军 任 莹(1.武汉轻工大学动物营养与饲料科学湖北省重点实验室,武汉430023;2.动物营养与饲料安全湖北省协同创新中心,武汉430023)
猪可代谢营养评定新体系研究进展
赵胜军1,2任 莹1,2
(1.武汉轻工大学动物营养与饲料科学湖北省重点实验室,武汉430023;2.动物营养与饲料安全湖北省协同创新中心,武汉430023)
在动物体内,葡萄糖具有重要的生理作用。糖代谢在动物能量代谢中处于重要的核心地位。现行营养评价指标体系大多建立在能量指标基础上,忽视了糖代谢在能量代谢中的核心地位,割裂了能量代谢和物质代谢的紧密联系,具有很强的表观性和局限性,有待于进一步发展。本文介绍了以代谢葡萄糖(MG)、代谢蛋白质(MP)和代谢生脂物质(MLS)为基础的物质指标体系的理论框架及其在猪的营养评价上的研究进展,为构建猪可代谢营养评定新体系进行了初步的探索。
猪;可代谢营养;评定体系
在动物生产中,所有的饲养标准都由两部分组成:一部分是对饲料养分含量的估计,可称之为“饲料营养价值评定”;另一部分是为使动物达到一定的生产水平所需的养分量,可称之为“营养需要量”。而这两者的量是以一系列的“营养素”为指标来表征的,我们称之为“营养价值评定体系”,简称“营养评定体系”。理论上讲,一套完善的营养评定体系是可以比较准确地预测动物生产性能的。但由于现行的猪营养评价指标体系大多建立在能量指标基础上,缺乏葡萄糖的评定指标,忽视了糖代谢在能量代谢中的核心地位,从而割裂了能量代谢和物质代谢的紧密联系,使其具有很强的表观性和局限性,在生产中大量使用粗饲料或在仔猪和母猪采食量较低时,预测生产性能的准确性较差。为此,本文在重新回顾糖代谢及其重要生理作用的基础上,介绍了以代谢葡萄糖(MG)、代谢蛋白质(MP)和代谢生脂物质(MLS)为基础的物质指标体系的理论框架,并介绍了为构建猪可代谢营养评定新体系进行的初步研究,希望能唤起学界同仁对猪可代谢营养评定新体系研究的兴趣和热情。
1 糖代谢及其重要生理作用
1.1 葡萄糖的生理作用
葡萄糖,化学式为C6H12O6,在生物学研究中具有重要地位,是活细胞的能量来源和新陈代谢中间产物,参与体内重要代谢过程。葡萄糖是中枢神经系统细胞,尤其是脑细胞的重要能量来源[1]。在免疫上,葡萄糖能够显著增加免疫功能[2],高浓度的葡萄糖能够影响直肠癌肿瘤细胞分泌免疫抑制物质[3]。在肌肉中,肌糖原分解为葡萄糖来满足肌肉运动或氧供应不足时机体所需的能量,并可以调控氨基酸(AA)转运以及可以作为仔猪骨骼肌蛋白质合成的重要信号分子[4-5]。在脂肪合成上,葡萄糖可以通过磷酸戊糖途径合成脂肪代谢中所必需的还原型辅酶Ⅱ(NADPH),脂肪组织会因葡萄糖的摄取增多而合成增加[6]。在肠道和肝脏中,小肠吸收葡萄糖是动物生长过程中所必不可少的一步,也是影响淀粉等饲料营养物质在动物小肠消化的重要因素[7-9]。经肠道吸收的葡萄糖可以合成肝糖原储存于肝脏中。肝糖原会影响机体对毒物或其他化学物质的防御及解毒作用。研究表明,机体肝糖原水平较高时,对疾病的抵抗力较强[10]。在生殖和泌乳方面,葡萄糖的充足供给是卵母细胞代谢的基本要求,也是胚胎早期发育的能源。在精子和卵细胞融合过程中,葡萄糖的代谢也是必需的[11]。对于繁殖母畜,血液中的葡萄糖也影响着乳腺的发育,并且是合成乳糖的唯一前体物。赵珂等[12]研究表明,增加葡萄糖的供应,可以通过增加乳糖合成底物的浓度,促进乳糖合成,而葡萄糖的酵解和磷酸戊糖途径的代谢也随之增加,进而促进细胞能量的合成,最终提高乳腺中乳脂合成量。
1.2 体内葡萄糖的来源
动物体内葡萄糖的来源主要有2个方面,即外源葡萄糖和内源葡萄糖。外源葡萄糖是由小肠消化、吸收的葡萄糖,主要由淀粉、纤维等水解后产生。内源葡萄糖即是由非糖物质(如乳酸、乙酸、丙酸等)在肝脏转化而来,这个过程称为糖的异生作用[13]。反刍动物体内的葡萄糖主要来自于瘤胃丙酸糖异生产生的内源葡萄糖。在猪和禽等单胃家畜大肠中,部分在肠前段未被消化吸收的营养物质(主要是碳水化合物)由微生物发酵分解,也可生成挥发性脂肪酸,其中的丙酸、乳酸也可在肝脏经糖异生转化为葡萄糖。Varel等[14]的研究表明,可溶性纤维在大肠中经微生物发酵作用,可以产生大量的挥发性脂肪酸,而这些挥发性脂肪酸能够提供猪维持需要能量的30%。也有研究表明,饲粮中抗性淀粉的比例越大,大肠中挥发性脂肪酸产生的量就越多[15]。
1.3 葡萄糖在体内的代谢
饲料来源的碳水化合物在消化道经消化酶消化后,绝大多数是以葡萄糖形式被吸收。对于那些酶解产物不是葡萄糖的也要经过肠道黏膜酶类的作用首先改造成为葡萄糖后,再被吸收进入肝脏。这种转化形式是单胃动物对饲料来源的碳水化合物的主要转化形式。进入肝脏的葡萄糖在葡萄糖激酶作用下磷酸化成为6-磷酸葡萄糖,后者再转变成1-磷酸葡萄糖,为葡萄糖与糖原分子连接做准备。1-磷酸葡萄糖与尿苷三磷酸(UTP)反应生成尿苷二磷酸葡萄糖(UDPG)和焦磷酸。由于焦磷酸在肝脏内迅速被焦磷酸酶水解,使反应向着糖原合成方向进行。其中,UDPG可看作为“活性葡萄糖”,充作葡萄糖的供体。
在动物饥饿状态下,糖异生是肝脏供应血糖的主要途径。大量饲喂粗料或处于绝食状态下的绵羊肝脏内产生的葡萄糖占整个机体葡萄糖周转量的85%~90%[16]。而肝脏糖异生作用的主要前体物是由消化道层次进入肝脏的丙酸,它占葡萄糖合成量的近1/2。在肝脏用于糖异生作用的物质中乳酸只占15%,氨基酸和甘油等也占一定比例。在绝食状态下,丙酸和乳酸用于糖异生作用的比例大大下降;而甘油则成为糖异生作用的主要前体物(占到40%)[17]。单胃家畜大肠发酵产生的丙酸和乳酸,也可在肝脏中经糖异生转化为葡萄糖,但所产生的葡萄糖占整个机体葡萄糖周转量的比例目前还尚未见报道。
肝糖原分解是指肝糖原分解成葡萄糖的过程。由肝糖原分解而来的6-磷酸葡萄糖,除了水解成葡萄糖而释放出之外,也可循酵解途径或磷酸戊糖途径进行代谢。当动物遭受饥饿时,后2条代谢途径均被抑制,肝糖原则绝大部分分解成葡萄糖释放入血液,运出肝脏外。在饱食状态下,进入肝脏的大量葡萄糖被合成糖原贮存起来。肝脏中绝大多数葡萄糖的分解代谢的主要形式是通过有氧氧化生成ATP而供能的。少部分葡萄糖也可经磷酸戊糖途径代谢生成磷酸核糖、NADPH和CO2。其主要生理意义是为核酸的生物合成提供核糖和提供NADPH作为供氢体参与多种代谢反应,而不是生成ATP。
1.4 糖代谢与脂肪和蛋白质代谢的关系
在生物体内,糖类、脂肪和蛋白质这3类物质的代谢是同时进行的,它们之间既相互联系,又相互制约,形成一个协调统一的整体。糖类在分解过程中产生的中间产物如丙酮酸,可以通过转氨基作用产生某些非必需氨基酸,同时也可以合成甘油和脂肪酸,形成脂肪。但由于糖类分解时不能产生与必需氨基酸相对应的中间产物,因而糖类不能转化成必需氨基酸。几乎所有组成蛋白质的天然氨基酸都可以转变成糖类,但只有少量的脂肪可以转变为糖类。因为脂肪分解产生的甘油和脂肪酸中,脂肪酸一般不能转变为糖类。
糖类、脂肪和蛋白质之间的转化是有条件的。只有在糖类供给充足的情况下,糖类才有可能大量转化成脂肪储存。此时,机体以葡萄糖供能为主,脂肪和蛋白质的分解供能减少。当体内葡萄糖供应不足时或饥饿时,体内的糖原因消耗减少,就会动用体内贮存的脂肪分解供能。若长期饥饿,体内脂肪分解大大加快,氧化生成的酮体过多,就会引起酮血症;糖的异生作用加强,动用体蛋白质转变为糖,以维持体内血糖含量的稳定,从而导致蛋白质利用率降低。
可见,糖代谢在能量代谢中处于中心地位,脂肪和蛋白质代谢处于次要的补充地位,三者的关系如图1所示。有报道称,在牛的饲粮中添加蔗糖可以减少体内氨基酸的生糖作用,同时减少尿素分解[18]。也有研究表明,增加小肠的可消化蛋白质数量能提高小肠淀粉消化率和门静脉血液中的葡萄糖净吸收[19]。因此,在研究动物体内蛋白质和脂肪代谢时,必须要同时关注葡萄糖代谢,并进行整体优化思考。

图1 糖类、脂肪和蛋白质代谢关系示意图
2 动物可代谢营养评定体系的提出及其意义
目前,几乎所有的现行营养评定体系都是将能量作为一个单独的营养素,并以此为基础确定蛋白质、纤维、钙、磷等各种营养素的需要量(图2)。事实上,无论是消化能、代谢能还是净能,都是以碳水化合物、粗蛋白质和粗脂肪的量乘以相应能值计算的总和。例如,根据饲料的可消化养分推算饲料消化能的公式为:
消化能(kJ/kg)=19.31X1+42.1X2+18.5X3+17.0X4。
式中:X1、X2、X3、X4分别代表每千克饲料中所含可消化粗蛋白质、可消化粗脂肪、可消化粗纤维和可消化无氮浸出物的克数;X1、X2、X3、X4前数字为各种物质的平均能值。
饲料的能量代表了碳水化合物、粗蛋白质和粗脂肪氧化后产生能量的潜力。但在通常情况下,动物体内所需的能量主要是由糖代谢提供的,脂肪和蛋白质则更多的是以物质形式沉积下来。刘飞飞[20]研究了消化能相同、碳水化合物/油脂比不同的饲粮对断奶仔猪生长性能的影响,发现碳水化合物/油脂比不同的饲粮的利用率存在显著差异。可见,能量指标是一个很笼统的指标,存在很大的表观性。现行营养评定体系以能量指标为基础,缺乏对机体葡萄糖供应情况的评估,有可能导致动物的葡萄糖供应不足,蛋白质分解供能增加,从而使其饲料利用率降低。

MP:代谢蛋白质metabolizable protein;DE:消化能 digestible energy;ME:代谢能metabolizable energy:Lys:赖氨酸 lysine;Met:蛋氨酸 methionine;Thr:苏氨酸 threonine;Trp:色氨酸tryptophan。
图2 现行营养评定体系的主要指标
Fig.2 The key indicators in current nutrition evaluating system
在动物体内,能量代谢和物质代谢是一个过程的2个方面。物质代谢是指物质在体内的消化、吸收、转运、分解等与生理有关的化学过程。这些物质在消化系统内需经一系列消化酶的分解,成为比较简单的有机物,才能被小肠所吸收。所以,物质代谢也即碳水化合物、蛋白质和脂肪三大养分的代谢。而能量代谢,主要是指动物体内的三大养分代谢过程中产生的能量利用与转化。可见,物质代谢与能量代谢是紧密联系着的。现行的营养评定体系把碳水化合物、蛋白质和脂肪换算成了能量,忽视了这些能量载体物质在供能作用上的差异,将能量代谢和物质代谢人为地割裂开来。由于能量是由3种物质所含能值换算来的,不是单一的物质,所以用能量指标只能对养分利用的结果进行描述,但却很难对能量代谢过程做到有针对性的调控。此外,现行营养评定体系还存在测定方法困难,难以得到实时数据;测定成本高,难以获得大量数据等缺陷。
针对现行营养评定体系存在的表观性问题,为了突出糖类在能量供应上的中心地位,1996年卢德勋[21]将反刍动物瘤胃产生的丙酸和小肠提供的葡萄糖所含的C3+C6能整合为一个整体性的技术指标,提出了代谢葡萄糖(metabolizable glucose,MG)的概念:饲料或饲粮中经动物消化、吸收后,可以给动物本身代谢提供的可利用的葡萄糖总量。孙海洲等[22]的研究表明,在1.1倍维持需要的饲养水平下,体重40 kg绵羊每日MG的最高水平为131.16 g或8.24 g/kg W0.75,MG能占代谢能的比例为32.83%。王玲[23]在1.2倍维持需要的饲养水平下,对体重23 kg的内蒙古白绒山羊生长肥育羊(处于非产绒期)的MG适宜水平进行了研究。该研究表明,绒山羊每日适宜的MG水平为44.13 g或4.39 g/kg W0.75,MG能占代谢能的比例应在12.28%以上。这些初步研究均说明,在动物饲粮内存在一个适宜的MG能占代谢能的比例,保持适宜的比例可以显著地提高能量利用率。此后,经过一系列的研究,初步建立了MG的技术体系,为反刍动物葡萄糖营养调控技术措施的提出奠定了基础。
在MG研究的基础上,为了进一步研究糖代谢对蛋白质代谢的影响,苏鹏程[24]应用MG和MP 2个指标研究了不同MG水平饲粮条件下内蒙古白绒山羊体内蛋白质(氨基酸)分配规律。研究表明,MG与小肠理想氨基酸模型下的MP之间存在适宜比例,生绒适宜MG/MP为1.06,整体适宜MG/MP为1.64。
为了体现脂肪在能量和物质代谢上的作用,2004年卢德勋[25]又将ME中的乙酸和丁酸所含的C2+C4能进行整合,提出了MLS的概念:饲料或饲粮经动物消化吸收后,可以给动物本身提供的,可利用的生脂物质(折合成乙酸)总量,包括乙酸、丁酸和脂肪3个部分。此后,杜瑞平[26]根据MLS这一新概念,测定了绵羊基础饲粮的MLS为135.69 g/d,(C2+C4)能占饲粮ME的89.24%;另外,还计算出基础饲粮的MG为43.83 g/d,(C3+C6)能占饲粮ME的11.30%,MLS/MG为3.10。这些数据为采用(C3+C6)能与(C2+C4)能在反刍动物饲粮ME中的调控及反刍动物能量调控整体优化提供了技术依据。
为了系统地阐述上述理论,卢德勋[27]提出了“能量载体物质”(energy-yielding substances)的概念。“能量载体物质”是指:饲料或动物机体内那些经过代谢可以为动物机体提供能量的物质。在动物体内,能量载体物质有2种前途:作为“燃料”,为动物机体生命活动提供ATP;作为动物机体内合成代谢的前体物。能量载体物质主要包括生糖物质、生氨基物质和生脂物质(图3)。以“能量载体物质”指标为基础,可构建由“MG”、“MP”和“MLS”组成的可代谢营养评定新体系(图4)。在应用新的评定体系调控动物营养平衡时,首先要保证能量载体物质内部的MG、MP和MLS的结构平衡,同时也要注意MG、MP和MLS内部组成成分的平衡,然后再考虑矿物质元素和维生素的平衡。

图3 能量载体物质的组成
可见,以能量载体物质作为营养评定新体系,能将能量代谢和物质代谢有机地结合起来,实现了能量代谢指标与蛋白质和脂肪代谢指标的完全整合,弥补了现行营养评定体系的不足,为能量代谢和物质代谢整体调控创造了条件,并有望使营养学在理论和应用上产生重大的突破。
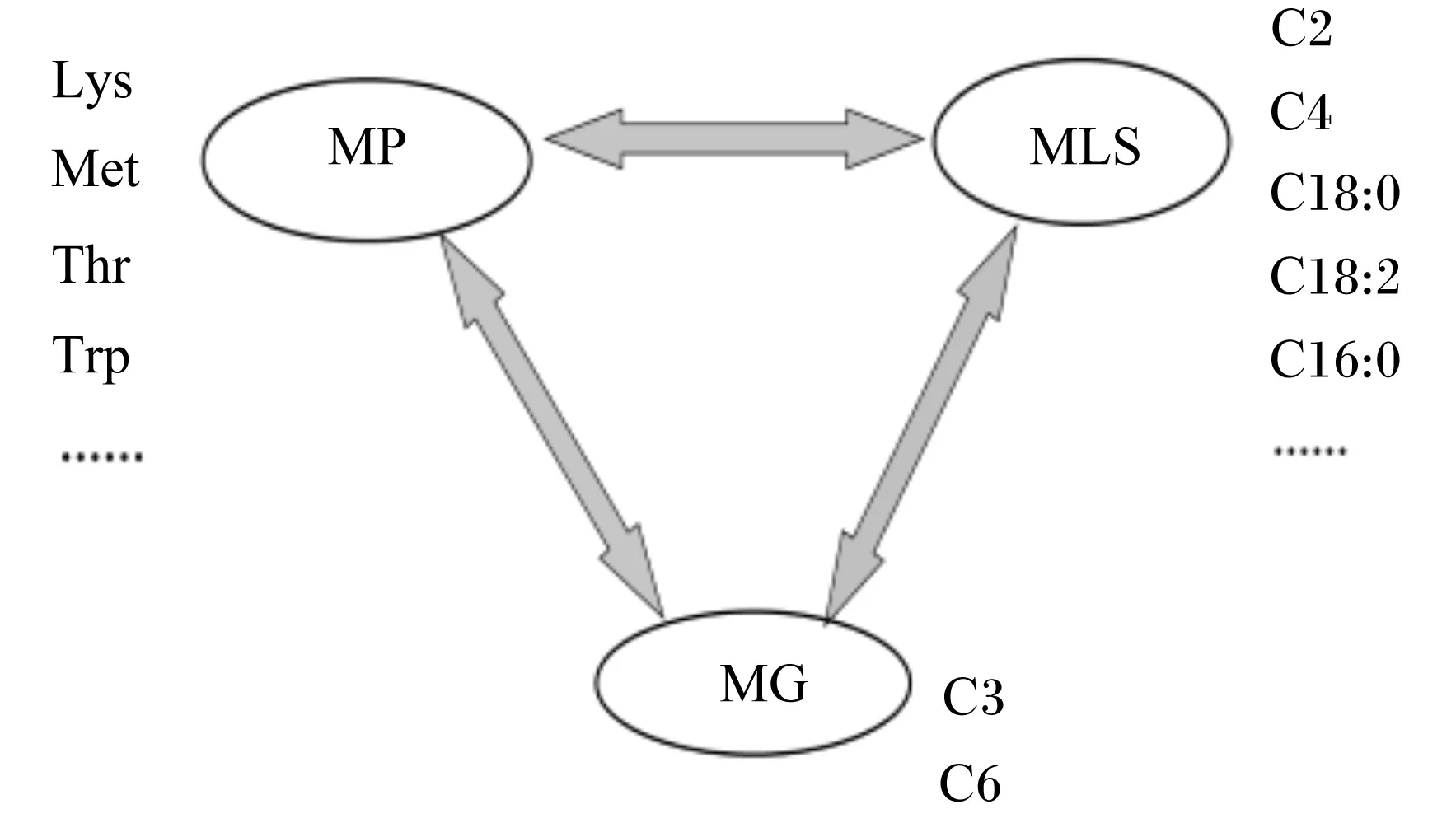
MP:代谢蛋白质 metabolizable protein;MG:代谢葡萄糖 metabolizable glucose;DLS:代谢生脂物质 metabolizable lipogenic substances;Lys:赖氨酸 lysine;Met:蛋氨酸 methionine;Thr:苏氨酸 threonine;Trp:色氨酸 tryptophan。
图4 可代谢营养评定新体系的主要指标
Fig.4 The key indicators in new evaluating system of metabolizable nutrition
3 猪可代谢营养评定体系的构建和进展
目前,猪的饲粮配合技术都是围绕着猪的饲养标准(或营养需要)展开的。现行的猪饲养标准同样是建立在能量指标基础上的,缺乏评定可代谢葡萄糖的指标。因此,人们在做配方时只能计算能量和蛋白质供需平衡,很难考虑猪的可代谢葡萄糖需要是否得到满足,容易造成猪体内葡萄糖供应缺乏,致使机体动员脂肪和蛋白质进行糖异生,从而导致了饲料利用率的降低。严重时,甚至会使猪体代谢紊乱,产生酮血症和尿酸中毒等症状。由此可见,采用卢德勋先生提出的“能量载体物质”指标,完善以能量指标为基础的现行营养评定体系,构建猪可代谢营养评定新体系,具有十分重要的理论和生产实践意义。
目前,猪的可代谢营养评定新体系的构建还处于起步阶段,主要集中在猪的MG营养方面的研究。2014年笔者根据卢德勋先生提出的MG的概念和猪的碳水化合物消化特点,建立了猪MG的计算公式[28]:
MG=IDSG+LFAG=0.9K1S+0.83K2Pr+0.87K3La。
式中:IDSG为外源葡萄糖,由淀粉在小肠提供的葡萄糖;LFAG为内源葡萄糖,由大肠发酵产生的丙酸和乳酸转化形成的葡萄糖;S为小肠淀粉量(g/d);Pr为大肠发酵产生的丙酸量(mmol/d);La为大肠发酵产生的乳酸量(mmol/d);K1为小肠淀粉消化率;K2、K3分别为丙酸和乳酸的吸收率;0.9、0.83、0.87分别为淀粉、丙酸及乳酸与葡萄糖的转化系数。
在此基础上,方勇[28]通过给断奶仔猪安装回肠瘘管来测定小肠淀粉消化率,建立了体内法测定断奶仔猪外源MG的方法,并测定了断奶仔猪4种饲粮的外源MG值分别为254.52、315.04、451.21、490.27 g/kg;此外,通过离体消化法模拟猪胃-肠道对4种饲粮进行体外消化,得到了体外法估算外源MG的公式。
张丹丹[29]通过给断奶仔猪安装颈动脉、门静脉和肠系膜血管瘘管,采用动-静脉浓度差法,建立了内源MG的测定方法,并结合方勇[28]的外源MG的测定方法,测得断奶仔猪4种饲粮的MG水平分别为239.62、280.68、400.79和454.35 g/kg;同时,又根据估算的饲粮MG水平与饲粮淀粉、粗纤维含量建立了MG的回归方程:Y=-196.44+12.13X1+23.18X2(Y表示MG水平,X1、X2分别表示饲粮中淀粉、粗纤维含量;R2=0.998 9,P=0.033),并在此基础上,通过比较不同MG水平饲粮对断奶仔猪生长性能的影响,最终确定了断奶仔猪适宜的MG需要量。
上述这些研究,为猪的葡萄糖营养调控技术的实用化奠定了基础,也为构建猪的可代谢营养评定新体系创造了良好的开端。今后,为了构建猪的可代谢营养评定新体系,还需开展猪的MP和MLS的研究,然后应用MG、MP和MLS研究猪不同生理阶段的营养需要量及平衡参数。同时,建立饲料的MG、MP和MLS体内和体外评定方法。最后,在MG、MP和MLS的基础上构建猪的可代谢营养评定新体系,建立新的猪饲养标准,指导养猪业的生产实践。当然,构建猪的可代谢营养评定新体系的研究工作,是事关我国猪营养学和猪生产学发展的重大战略举措,工作量大、任务重,并非某个个人或研究小组所能完成的,还需引起学界相关领导和部门的重视,在国家层面开展系统的研究和推进,以有效开展工作,从而尽快使中国猪的营养研究和应用走在世界前列。
[1] 张雪竹,刘存志,于建春,等.脑组织葡萄糖代谢的研究方法[J].国际脑血管病杂志,2006,14(10):785-788.
[2] 付辰炜.氨基葡萄糖及壳寡糖对小鼠免疫功能的影响[D].硕士学位论文.青岛:中国海洋大学,2006.
[3] 周慧敏.不同浓度的葡萄糖和胰岛素对Colon26肿瘤细胞分泌免疫抑制物质的影响及其分子机制的研究[D].博士学位论文.石家庄:河北医科大学,2009.
[4] JEYAPALAN A S,ORELLANA R A,SURYAWAN A,et al.Glucose stimulates protein synthesis in skeletal muscle of neonatal pigs through an AMPK- and mTOR-independent process[J].American Journal of Physiology:Endocrinology and Metabolism,2007,293(2):E595-E603.
[5] ROOS S,LAGERLÖF O,WENNERGREN M,et al.Regulation of amino acid transporters by glucose and growth factors in cultured primary human trophoblast cells is mediated by mTOR signaling[J].American Journal of Physiology:Cell Physiology,2009,297(3):C723-C731.
[6] 汪晓芬.大鼠追赶生长早期脂肪组织和骨骼肌葡萄糖代谢差异及其机制的研究[D].硕士学位论文.武汉:华中科技大学,2008.
[7] 孙海洲,卢德勋.十二指肠灌注不同水平的玉米淀粉对生长肥育绵羊小肠内葡萄糖消化及吸收的影响[J].动物营养学报,1999,11(增刊):173-185.
[8] 王文娟,汪水平,左福元.反刍动物淀粉消化与葡萄糖吸收研究进展[J].西北农林科技大学学报:自然科学版,2008,36(5):27-34.
[9] 宾石玉.日粮淀粉来源对断奶仔猪生产性能、小肠淀粉消化和内脏组织蛋白质合成的影响[D].博士学位论文.雅安:四川农业大学.2005.
[10] 赵文卓.碳水化合物的重要作用[J].科技创新导报,2012(31):223.
[11] 胡承阅.葡萄糖参与小鼠精子与卵细胞的融合[J].国外医学:计划生育分册,1997(2):107.
[12] 赵珂,刘红云,刘建新.泌乳奶牛乳腺葡萄糖吸收、代谢及其调控研究进展[J].中国奶牛,2009(8):26-30.
[13] 张军,黄瑞林,李铁军,等.葡萄糖代谢调控[J].广西农业生物科学,2006,25(1):86-90.
[14] VAREL V H,YEN J T.Microbial perspective on fiber utilization by swine[J].Journal of Animal Science,1997,75(10):2715-2722.
[15] HEDEMANN M S,KNUDSEN B K E.Resistant starch for weaning pigs-effect on concentration of short chain fatty acids in digesta and intestinal morphology[J].Livestock Science,2007,108(1/2/3):175-177.
[16] BERGMAN E N,KATZ M L,KAUFMAN C F.Quantitative aspects of hepatic and portal glucose metabolism and turnover in sheep[J].The American Journal of Physiology,1970,219(3):785-793.
[17] BERGMAN E N.Energy contributions of volatile fatty acids from the gastrointestinal tract in various species[J].Physiological Reviews,1990,70(2):567-590.
[18] SANNES R A,MESSMAN M A,VAGNONI D B.Form of rumen-degradable carbohydrate and nitrogen on microbial protein synthesis and protein efficiency of dairy cows[J].Journal of Dairy Science,2002,85(4):900-908.
[19] 邓代君,孟庆翔.日粮中碳水化合物和含氮化合物对奶牛泌乳性能的影响[J].饲料工业,2005,26(19):41-43.
[20] 刘飞飞.日粮能源结构对仔猪氮素利用的影响及机制研究[D].博士学位论文.长春:吉林农业大学,2015.
[21] 卢德勋.在动物营养学研究领域应用系统科学的初步探索[C]//动物营养研究进展.北京:中国农业大学出版社,1996.
[22] 孙海洲.生长肥育羊葡萄糖营养整体优化规律的研究[D].博士学位论文.呼和浩特:内蒙古农业大学,1999.
[23] 王玲.内蒙古白绒山羊适宜代谢葡萄糖水平的评定[D].硕士学位论文.呼和浩特:内蒙古农业大学,2003.
[24] 苏鹏程.不同代谢葡萄糖水平日粮条件下白绒山羊体内蛋白质(氨基酸)分配规律的研究[D].博士学位论文.呼和浩特:内蒙古农业大学,2004.
[25] 卢德勋.系统动物营养学导论[M].北京:中国农业出版社,2004.
[26] 杜瑞平.绵羊瘤胃乙酸和丁酸的产生、吸收和利用规律及可代谢生脂物质(MLS)的测定[D].硕士学位论文.呼和浩特:内蒙古农业大学,2006.
[27] 卢德勋.新版系统动物营养学导论[M].北京:中国农业出版社,2016.
[28] 方勇.断奶仔猪外源代谢葡萄糖的评定[D].硕士学位论文.武汉:武汉轻工大学,2015.
[29] 张丹丹.断奶仔猪代谢葡萄糖事宜需要量的研究[D].硕士学位论文.武汉:武汉轻工大学,2016.
Author, associate professor, E-mail: zhaoshengjun1974@163.com
(责任编辑 菅景颖)
Research Development of New Evaluating System of Metabolizable Nutrition in Swine
ZHAO Shengjun1,2REN Ying1,2
(1.HubeiKeyLaboratoryofAnimalNutritionandFeedScience,WuhanPolytechnicUniversity,Wuhan430023,China; 2.HubeiCollaborativeInnovationCenterforAnimalNutritionandFeedSafety,Wuhan430023,China)
Glucose plays an important role in animal physiology, as important as the carbohydrate metabolism in the energy metabolism of animals. The current nutrition evaluating systems are apparent and limited, which set in the basis of energy indication, neglect the important position of carbohydrate metabolism, and separate the interaction between energy metabolism and substance metabolism. The paper has introduced the index system of material and its research development in swine on the basis of metabolizable glucose (MG), metabolizable protein (MP) and metabolizable lipogenic substances (MLS), which has preliminarily explored the evaluating system of metabolizable nutrition.[ChineseJournalofAnimalNutrition, 2017, 29(5):1455-1461]
swine; metabolizable nutrition; evaluating system
10.3969/j.issn.1006-267x.2017.05.001
2016-10-31
赵胜军(1974—),男,蒙古族,内蒙古通辽人,副教授,博士,主要从事动物营养与代谢调控研究。E-mail: zhaoshengjun1974@163.com
S816.1
A
1006-267X(2017)05-1455-07
