氮、磷施肥对马尾松幼苗生长和叶片化学计量特征的影响
2017-05-02王满堂靳冰洁郑媛李曼程栋梁
王满堂 靳冰洁 郑媛 李曼 程栋梁
摘要:为了探究马尾松对氮、磷养分的响应机制,以一年生马尾松(Pinus massoniana)幼苗为研究对象,通过盆栽施肥试验,研究氮、磷施肥对马尾松幼苗生长的影响,以期为马尾松苗木培育、养分管理和幼林施肥提供理论依据。结果表明:不同施肥处理,马尾松苗木叶片氮含量范围在(10.13±0.97)~(19.88±1.70) mg/g,磷含量范围在(0.30±0.10)~(1.18±0.13) mg/g,氮磷比范围在(10.53±1.79)~(50.07±17.52)之间。施氮处理下,马尾松苗木的地径生长速率与株高生长速率呈极显著正相关(P<0.01);叶片氮含量与苗木株高生长速率呈极显著正相关关系(P<001)。施磷处理下,马尾松苗木的地径生长速率与株高生长速率呈极显著正相关(P<0.01),磷含量与苗木株高生长速率呈极显著正相关关系(P<0.01)。
关键词:马尾松幼苗;氮肥;磷肥;生长速率;叶片化学计量
中图分类号:S791.248.01文献标志码: A
文章编号:1002-1302(2017)03-0104-06
[HJ1.4mm]
收稿日期:2015-12-22
基金项目:国家自然科学基金(编号:31170374、31370589、31170596);福建省教育厅新世纪优秀人才支持计划(编号:JA12055);福建省杰出青年基金(编号:2013J06009);山东省自然科学基金(编号:ZR2013CL027)。
作者简介:王满堂(1980—),男,山东省枣庄人,博士,副教授,主要从事植物生态学、理論生态学研究。Tel:(0632)3786019;E-mail:wangmantang@aliyun.com。
通信作者:程栋梁,博士,教授,主要从事植物生理生态学研究。E-mail:chengdl02@aliyun.com。[HJ]
生态化学计量学是研究生态系统能量平衡和多重化学元素平衡的科学[1],主要研究生命有机体主要组成元素间的关系,特别是碳、氮和磷等重要元素间的计量关系[1]。目前,生态化学计量学的研究主要集中在种群、群落结构动态、生物养分限制、营养级动态、生物地球化学循环、生态系统养分循环与供需平衡等领域,并取得了大量研究成果[1-7]。
氮、磷是陆地生态系统中重要的营养元素,也是限制性生态因子,能够影响植物个体的生长状态、群落的动态变化乃至整个生态系统的生产力[8-10]。关于氮、磷关系的假说主要有2个,一个是内稳态假说,另一个是生长速率假说(growth rate hypothesis,简称GRH)。生长速率假说认为生物体在快速生长阶段需要投入更多富磷rRNA来支持蛋白质的合成。rRNA是植物的主要磷库,rRNA含量的增加将导致细胞中磷浓度的增加,从而使植物表现出较低的氮磷比。
马尾松是主要分布于亚热带地区的常绿乔木,是我国南方地区的主要造林树种,木材用途广泛,兼具经济价值和观赏价值。本研究以一年生马尾松幼苗为研究对象,采用人工受控试验方法,设置不同浓度梯度的氮、磷施肥试验,研究氮、磷营养对马尾松幼苗生物量的分配及氮、磷养分分配的影响,探讨马尾松苗木对氮、磷营养的需求特性和吸收利用机制,明确不同养分供给条件下马尾松生长与养分分配的基本规律,以期揭示不同氮、磷浓度下马尾松幼苗的生长速率及叶片化学计量特征,为马尾松苗木培育、养分管理和幼林施肥提供基础理论依据。
1材料与方法
1.1试验区概况
[JP3]福建师范大学长安山公园气象站苗木试验区(26°05′41″N,[JP]118°08′47″E),海拔约为100 m,属于亚热带季风气候,温暖湿润,雨量充沛,霜少无雪,夏长冬短。年均降水量为 900~2 100 mm,多集中于3—10月份,年均温度约为 19.6 ℃,1月最冷月均温10.5 ℃,7月最热月均温28.6 ℃。年均日照时间为1 840~1 980 h,无霜期为326 d,≥10 ℃年积温6 100~6 300 ℃。原始土壤为红壤,其中土壤氮含量为0.518 mg/g,磷含量为5.417 mg/kg,pH值为4.78,呈酸性,土壤有机质含量为0.40%,含水率为11.17%。根据1992年中国第二次土壤普查推荐土壤养分定级标准,本试验区土壤有机质匮乏,处于中下水平。
1.2试验材料
2013年4月,从福建省南平市顺昌县林业科技推广中心选取生长良好的一年生马尾松幼苗带回福建师范大学长安山公园气象站苗木试验区并移入实验盆内进行预培植,其中花盆为底径12 cm的塑料盆,每盆基质土质量为3 kg。5~6个月后,幼苗适应试验区的自然环境,选取成活并长势良好的苗木,测定其初始株高均值为(15.53±1.61) cm,初始地径均值为(2.52±0.18) mm。
1.3试验设计
基于试验区土壤营养状况,氮、磷施肥处理各设定4个梯度N1、N2、N3、N4及P1、P2、P3、P4,氮、磷施肥量分别是初始土壤的2、4、8、16倍,不同氮、磷浓度处理共用一组对照(CK),共9种处理。每个处理设置3个重复小区,每个重复小区有4盆。根据基质土壤的全氮、全磷浓度和施肥梯度计算应添加NH4NO3、NaH2PO4的量,将称好的NH4NO3、NaH2PO4溶解于水中,于每月月中晴朗的傍晚时均匀喷洒在盆栽容器土壤中,试验前控制水分以利于养分在盆土中的扩散。对照处理只浇水,不施加任何肥料。完成添加N、P处理后,分别于2014年3、7、11月初,定期采集苗木未受病虫害影响的叶片,按照不同的施肥处理分装、归类。将鲜叶用蒸馏水洗净,去除杂质,放置于105 ℃烘箱中杀青30 min,再置于65 ℃恒温下烘干至恒质量,磨碎过筛(筛孔尺寸0.147 mm)备用。
1.4指标测定
苗木地径和株高:分别于2014年3、7、11月初,用直尺和数显卡尺测量所有幼苗的苗高和地径。
叶片氮磷测定:利用碳氮元素分析仪(elemental analyzer vario EL Ⅲ)进行植物叶片氮含量的测定;利用连续流动分析仪(荷兰SKALAR)对植物叶片磷含量的测定。
1.5数据处理
生长速率的计算是以苗木的初始株高、地径为基准,计算苗木的平均相对生长速率,公式如下:
[JZ]R=(lnQ2-lnQ1)/(T2-T1)。
[JP3]式中:Q1为前一测量阶段的(T1)的地径、株高值,Q2为后一测量阶段(T2)的地径、株高值,ln为自然对数。株高的生长速率单位:cm/(cm·30 d),地径的生长速率单位:mm/(cm·30 d)。[JP]
采用Origin 9.0制作图表,利用SPSS 20.0进行单因素方差分析(One-way ANOVA),用LSD多重检验方法,在0.05水平上表示差异显著(P<0.05),检验各处理间的显著差异性。
2结果与分析
2.1不同氮、磷施肥处理下马尾松生长速率及动态变化
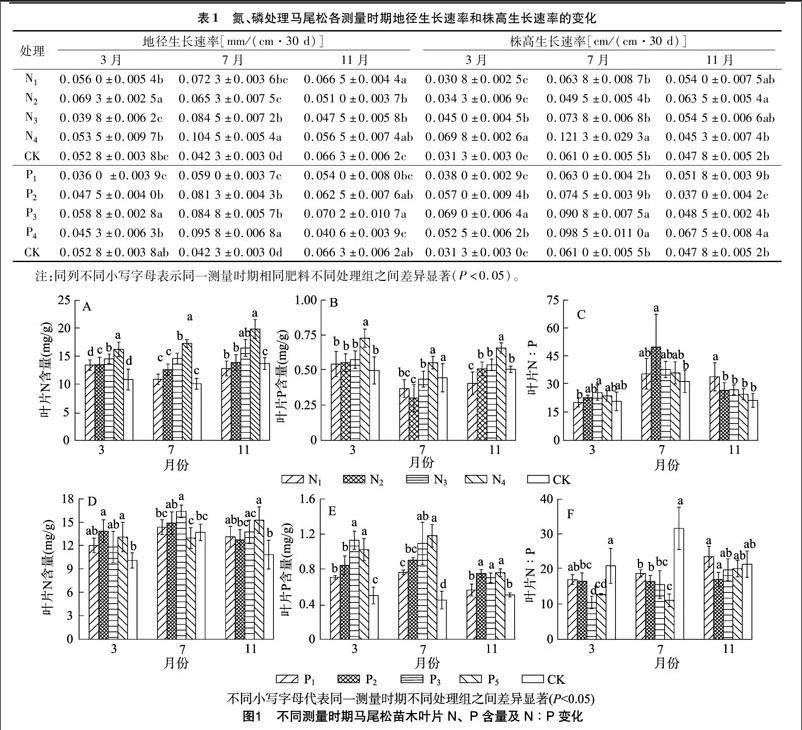
由表1可知,在施氮处理下,3、7、11月地径生长速率最大值分别为N2处理(0.069 3±0.002 5) mm/(cm·30 d),N4处理(0.104 5±0.005 4) mm/(cm·30 d),N1处理(0.066 5±0.004 4) mm/(cm·30 d);3、7、11月株高生长率最大值分别为N4处理(0.069 8±0.002 6) cm/(cm·30 d),N4处理(0.121 3±0.029 3) cm/(cm·30 d),N2处理(0.063 5±0.005 4) cm/(cm·30 d)。在施磷处理下,3、7、11月地径生长速率最大值分别为P3处理(0.058 8±0.002 8) mm/(cm·30 d),P4处理(0.095 8±0.006 8) mm/(cm·30 d),P3处理(0.070 2± 0.010 7) mm/(cm·30 d);3、7、11月株高生长率最大值分别为P3处理(0.069 0±0.006 4) cm/(cm·30 d),P4处理(0.098 5±0011 0) cm/(cm·30 d),P4处理(0.067 5±0.008 4) cm/(cm·30 d)。[JP]
在不同测量时期,马尾松的地径生长速率和株高生长速率在氮(除N2处理)、磷施肥处理下呈先升高再降低的动态变化。其中7月处于苗木快速生長时期,高氮(初始土壤氮含量的16倍)、高磷(初始土壤磷含量的16倍)的施肥处理更能促进地径、株高的有效增长。
2.2氮、磷施肥对马尾松叶片氮、磷含量及氮磷比的影响
从图1可以看出,在生长初期(3月初)施氮肥(N1处理除外)显著增加了马尾松叶片的氮含量(P<0.05),而仅在高氮(初始土壤氮含量的16倍)处理下,叶片中磷的含量显著增加。在不同测量时期,随着供氮水平的提高,叶片氮累计含量增加。在生长初期(3月初)和生长末期(11月初),叶片磷累计含量呈逐渐上升趋势,而氮磷比呈先升后降的趋势。氮磷比均大于14,受磷限制(图1-A至图1-C)。
从图1-E可以看出,施磷肥(11月P1处理除外)显著增加了马尾松叶片的磷含量(P<0.05),不同浓度的施磷肥对叶片氮含量的影响不同(图1-D),而在3、7月不同处理间氮磷比随着叶片磷含量的增加而下降,11月,与对照相比,施磷肥处理对氮磷比影响差异不显著(P>0.05)(图1-F)。与对照相比,随着供磷水平的提高,叶片磷含量呈上升趋势(7月),而叶片氮含量、氮磷比变化复杂。
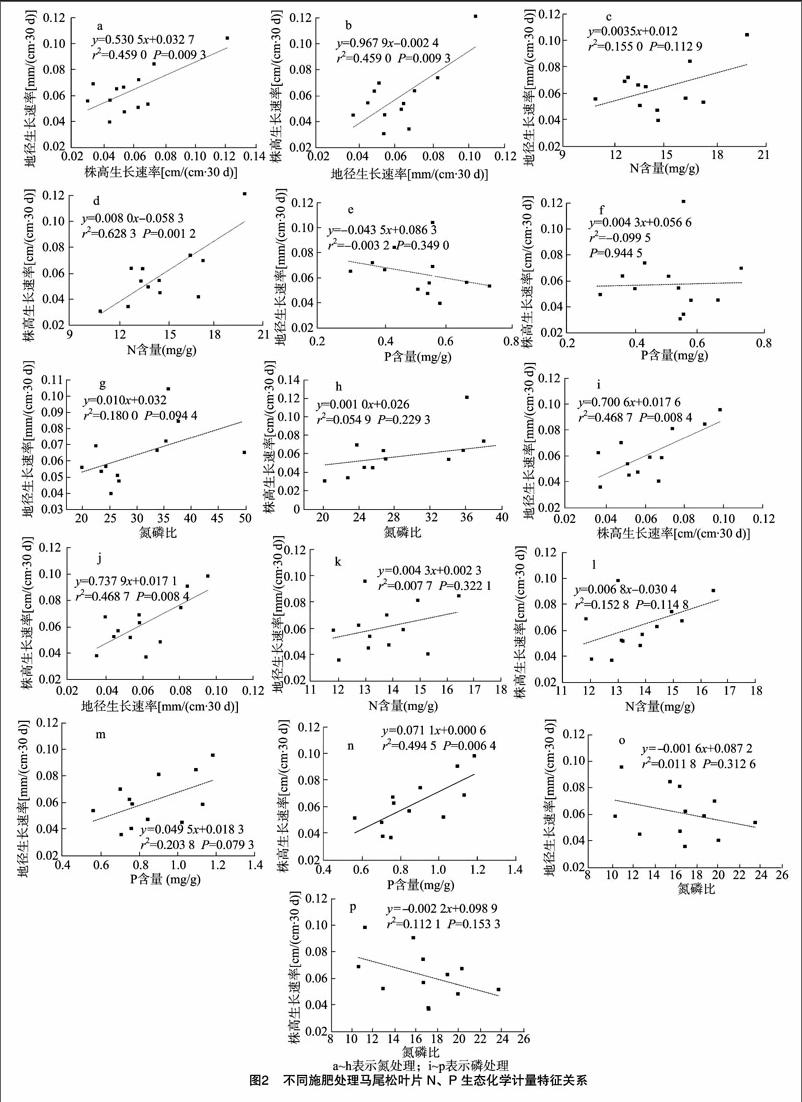
2.3马尾松叶片氮、磷及氮磷比与生长速率的关系
在不同的施肥处理下,马尾松叶片氮、磷及氮磷比与苗木的[CM(25]地径、株高生长速率具有一定的相关关系(图2)。在施氮处理下,马尾松苗木的地径生长速率与株高生长速率呈极显著正相关(P<0.01);叶片氮含量与苗木株高生长速率呈极显著正相关关系(P<0.01)。在施磷处理下,马尾松苗木的地径生长速率与株高生长速率呈极显著正相关关系(P<0.01);叶片磷含量与苗木株高生长速率呈极显著正相关关系(P<0.01)。在施肥处理中,叶片氮磷比与地径生长速率和株高生长速率没有表现出明显的相关关系。
3结论与讨论
3.1施肥对马尾松生长速率的影响
施肥直接改变了土壤中的养分格局,进而影响植物的氮、磷吸收机制,促使植物改变器官化学计量的分配、地上地下生物量的分配累积,从而更好地适应环境、竞争资源。植物地径、株高的生长速率直接反映了植物的养分吸收状况和有机质的累积,是评定植物生长的重要参考指标,增加陆地生态系统中的氮含量可以有效提高植物的光合能力和生态系统的初级生产力[11-13]。
施氮肥处理下,与对照相比,地径和株高的生长速率均有变化,同时在3—7月中,高氮(初始土壤氮含量的16倍)处理能够显著增加植物株高生长速率。一定范围内的氮素营养供给可以加强根部的磷酸酶活性,刺激植物根系的生长发育,并且对植物地上部分的生长作用更为显著[14-15]。此外,植物在受到氮胁迫环境下,地下部分分配到的碳水化合物高于地上部分,植物所吸收到的氮素优先用于满足地下部分的生长,导致地上部分的生长减弱[16]。因此马尾松苗木在高氮(初始土壤氮含量的16倍)处理下,能够促进苗木的快速生长。但是在植物不同的生长时期,不同处理组对苗木生长的促进作用不一,说明氮施肥能够促进植物生长,但是应采取不同的施肥策略, 在快速生长时期,以高氮(初始土壤氮含量的16倍)处理为主,而在非生长时期,中氮(初始土壤氮含量的8倍)处理为宜,这也与克热木·伊力等的部分研究结果[17]相类似。
施磷肥处理下,与对照相比,苗木的生长速率各有不同,且[CM(25]在整体上施磷肥对于促进苗木株高、地径的生长效果要稍[CM)][FL)]
[FK(W51][TPWMT2.tif][FK)]
逊于施氮肥。同时在非生长季时期,高磷(初始土壤磷含量的16倍)的施肥可能抑制了植物的生长,造成苗木的奢侈吸收,此时对于苗木的地径生长没有助益。植物吸收磷元素主要通过土壤途径,随着供磷水平增加而增加,但是植物难以直接利用土壤中不溶于水的无机盐和有机磷形式的磷元素,所以直接施加磷肥对于促进苗木的生长效果可能没有氮肥显著,因为磷元素易于被土壤固定[18-19]。王东光等通过对闽楠苗木施磷肥,研究发现供磷水平提高能使苗高和生物量先增加,再逐渐趋于稳定,而地径的增长则有波动,同时叶片氮、磷含量也会相应增加,表明磷元素的供给对于闽楠的生长有一个临界值[20]。本研究的结果与之相符。
3.2施肥对马尾松叶片氮、磷含量和氮磷比的影响
由图1可知,对于不同施肥处理,马尾松苗木叶片N含量范围在(10.13±0.97)~(19.88±1.70) mg/g,P含量范围在 (0.30±0.10)~(1.18±0.13) mg/g,N ∶[KG-*3]P范围在(10.53±1.79)~(50.07±17.52)之间。
在施氮肥处理下,叶片磷含量显著增长,这可能是因为随着氮的增加,植物将自身吸收的氮更多地作用于构建磷酸酶,刺激了根系的磷酸酶活性,从而吸收更多的磷使得叶片磷含量提高,说明氮可利用性的提高具有改善磷循环的作用[21-22]。Gusewell指出在植物水平上,短期肥力试验表明,当氮磷比小于10时,植物生物量受氮限制,当氮磷比大于20时,植物生物量受磷限制[23]。除了3月份低氮(初始土壤氮含量的2倍)处理外,其他处理在不同的测量时期,氮磷比大于20,受磷限制。本试验中原始土壤为红壤,磷素很容易被固定,且氮处理下没有施加外源磷肥,苗木生长受限制。在氮处理组内,3—11月,苗木的叶片氮含量都呈现逐渐上升趋势。施肥开始到次年3月,苗木处于萌芽前期,生物量较小,且处于冬季,气温低,对于所施加的养分不能完全吸收,而在快速生长期则氮含量增加,11月份生长缓慢期,叶片开始逐渐衰老,出现养分回流的现象,导致氮含量较之前减少[24]。生长初期(3月初)和生长末期(11月初)磷含量则呈现递增的趋势,因为3—7月是苗木速生期,植物快速生长需要更多的rRNA合成蛋白质用以支持生长,而rRNA是主要的磷贮藏库,因此对于磷的消耗较大[25]。杨阔等研究发现,植物叶片的氮磷比与温度之间具有显著关系[26]。从3月初和7月初可以发现,马尾松苗木氮磷比呈现先增加之后降低的趋势。由于3—7月气温增加,8—11月氣温降低,因此氮磷比呈现出不同的变化。在施磷肥处理组内,3月初和7月初的测量结果中发现叶片氮含量变化不同,而磷含量变化显著,导致氮磷比降低。这符合生长速率假说,快速生长的个体所需的磷元素相对更多,因此氮磷比会更低。
彭少麟研究表明,单独施用某一种元素,会明显增加该种元素在植物体内的含量,但是对于其他含量稳定的元素可能产生稀释效应,使其含量降低[27]。Baldwin等研究表明,叶片的养分含量与植物自身的生理特性以及生长季密切相关[28]。Orgeas等也发现植物的氮、磷浓度在不同生长季节差异明显[29]。本研究不同时期马尾松的氮、磷施肥各处理组之间叶片的氮、磷养分浓度也具有一定差异的结果与之相似。
3.3马尾松叶片氮、磷及氮磷比与生长速率的关系
植物的养分元素与其生长发育有着重要的联系,营养元素的供应限制着整个陆地生物圈的生产力。因此,植物叶片的氮、磷化学计量特征在很大程度上影响了植物的生长速率。有研究表明,植物叶片氮磷比值与植物生长速率相关[30-31]。也有研究发现,植物叶片的高氮、磷含量表示它们能够快速地吸收生境中的营养资源,通常这些植物种的生长速率较高;相反植物叶片氮、磷含量较低的物种侧重于保存资源,这些物种通常生长较慢,随着生长速率的增大,植物的氮、磷含量通常都会增加,而磷元素增加得更为显著,所以生长率高的植物常常有较低的氮磷比[32]。
Agren提出了植物相对生长速率假说模型,并通过试验进行了验证,他认为,植物体内氮元素的变化主要取决于磷元素,因为磷元素主要存在于rRNA中,氮元素主要存在于蛋白质中,而蛋白质的合成速率受rRNA浓度的控制,当植物体均衡生长的时候,植物个体的相对生长速率与其氮、磷含量呈正比,而与氮磷比呈反比[33]。
不同的施肥处理下,叶片的氮、磷含量与苗木的株高生长速率呈极显著的正相关关系,符合植物生长速率假说模型。然而,Matzek等研究了14种松树树苗的生长,发现叶片的蛋白质 ∶[KG-*3]RNA与生长速率呈显著负相关,而叶片的氮磷比与生长速率没有显著相关性[34],本试验中不同的施肥处理下,氮磷比与株高生长速率没有显著的线性关系,与其研究相一致。Loladze等通过理论模型和文献数据分析,研究认为N ∶[KG-*3]P的根源在于稳定的蛋白质 ∶[KG-*3]RNA[35]。因此,关于生长速率的调控机制尚须要进一步研究。
[HS2]参考文献:
[1]Reich P B,Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America,2004,101(30):11001-11006.
[2]Lu X,Mao Q,Gilliam F S,et al. Nitrogen deposition contributes to soil acidification in tropical ecosystems[J]. Global Change Biology,2014,20(12):3790-3801.
[3]Lal R. Soil carbon sequestration impacts on global climate change and food security[J]. Science,2004,304(5677):1623-1627.
[4]Schipper L A,Percival H J,Sparling G P. An approach for estimating when soils will reach maximum nitrogen storage[J]. Soil Use and Management,2004,20(3):281-286.
[5]Stevenson F J,Cole M A. Cycles of soils: carbon,nitrogen,phosphorus,sulfur,micronutrients[M]. New York:Wiley,1999.
[6]曾德慧,陳广生. 生态化学计量学:复杂生命系统奥秘探索[J]. 植物生态学报,2005,29(6):1007-1019.
[7]Reiners W A. Complementary models for ecosystems[J]. American Naturalist,1986,127(1):59-73.
[8]Kerkhoff A J,Fagan W F,Elser J J,et al. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants[J]. American Naturalist,2006,168(4):103-122.
[9]Lea R,Tierson W C,Bickelhaupt D H,et al. Differential foliar responses of northern hardwoods to fertilization[J]. Plant and Soil,1980,54(3):419-439.
[10]Crawford N M,Glass A D M. Molecular and physiological aspects of nitrate uptake in plants[J]. Trends in plant science,1998,3(10):389-395.
[11]Perring M P,Hedin L O,Levin S A,et al. Increased plant growth from nitrogen addition should conserve phosphorus in terrestrial ecosystems[J]. Proceedings of the National Academy of Sciences,2008,105(6):1971-1976.
[12]Granath G,Strengbom J,Breeuwer A,et al. Photosynthetic performance in Sphagnum transplanted along a latitudinal nitrogen deposition gradient[J]. Oecologia,2009,159(4):705-715.
[13]Fujita Y,Robroek B J M,Ruiter P C D,et al. Increased N affects P uptake of eight grassland species: the role of root surface phosphatase activity[J]. Oikos,2010,119(10):1665-1673.
[14]Kramer P J,Boyer J S. Water relations of plants and soils[M]. San Diego: Academic Press,1995.
[15]Wang J R,Hawkins C D B,Letchford T. Relative growth rate and biomass allocation of paper birch (Betula papyrifera) populations under different soil moisture and nutrient regimes[J]. Canadian Journal of Forest Research,2011,28(1):44-45.
[16]林文,李义珍. 施氮量及施肥法对水稻根系形态发育和地上部生长的影响[J]. 福建稻麦科技,1999,17(3):21-24.
[17]克热木·伊力,新居直祐. 不同氮素施用量对葡萄叶、枝、根碳水化合物含量的影响[J]. 新疆农业大学学报,2001,24(1):64-68.
[18]Barber S A. Soil nutrient bioavailability: a mechanistic approach [M]. New York: Wiley,1995.
[19]Holford I C R. Soil phosphorus: its measurement,and its uptake by plants[J]. Australian Journal of Soil Research,1997,35(2):227-240.
[20]王东光,尹光天,杨锦昌,等. 磷肥对闽楠苗木生长及叶片氮磷钾浓度的影响[J]. 南京林业大学学报(自然科学版),2014,38(3):40-44.
[21]Treseder K K,Vitousek P M. Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests[J]. Ecology,2001,82(4):946-954.
[22]Marklein A R,Houlton B Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems[J]. New Phytologist,2012,193(3):696-704.
[23]Gusewell S. N ∶[KG-*3]P ratios in terrestrial plants: variation and functional significant[J]. New Phytologist,2004,164(2):243-266.
[24]Lin Y M,Sternberg L,Silveria L D. Nitrogen and phosphorus dynamics and nutrient resorption of Rhizophora mangle leaves in south Florida,USA [J]. Bulletin of Marine Science,2007,80(1):159-169.
[25]程濱,赵永军,张文广,等. 生态化学计量学研究进展[J]. 生态学报,2010,30(6):1628-1637.
[26]杨阔,黄建辉,董丹,等. 青藏高原草地植物群落冠层叶片氮磷化学计量学分析[J]. 植物生态学报,2010,34(1):17-22.
[27]彭少麟. 热带亚热带恢复生态学研究与实践[M]. 北京: 科学出版社,2003.
[28]Baldwin D S,Rees G N,Mitchell A M,et al. The short-term effects of salinization on anaerobic nutrient cycling and microbial community structure in sediment from a freshwater wetland[J]. Wetlands the Journal of the Society of the Wetland Scientists,2009,26(2):455-464.
[29]Orgeas J,Ourcival J M,Bonin G. Seasonal and spatial patterns of foliar nutrients in cork oak (Quercus suber L.) growing on siliceous soils in Provence (France)[J]. Plant Ecology,2003,164(2):201-211.
[30]Niklas K J,Owens T,Reich P B,et al. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth[J]. Ecology Letters,2005,8(6):636-642.
[31]Niklas K J. Plant allometry,leaf nitrogen and phosphorus stoichiometry,and interspecific trends in annual growth rates[J]. Annals of Botany,2006,97(2):155-163.
[32]Elser J J,Obrien W J,Dobberfuhl D R,et al. The evolution of ecosystem processes: growth rate and elemental stoichiometry of a key herbivore in temperate and arctic habitats[J]. Journal of Evolutionary Biology,2000,13(5):845-853.
[33]Agren G I. The C ∶[KG-*3]N ∶[KG-*3] P stoichiometry of autotrophs-theory and observations[J]. Ecology Letters,2004,7(3):185-191.
[34]Matzek V,Vitousek P M. N ∶[KG-*3]P stoichiometry and protein: RNA ratios in vascular plants: an evaluation of the growth-rate hypothesis[J]. Ecology Letters,2009,12(8):765-771.
[35]Loladze I,Elser J J. The origins of the Redfield nitrogen-to-phosphorus ratio are in a homoeostatic protein-to-rRNA ratio[J]. Ecology Letters,2011,14(3):244-250.
