橘小实蝇成虫复眼结构及感光机制
2017-02-14邓淑桢赵晓峰曹凤勤陆永跃
刘 欢, 邓淑桢, 赵晓峰, 曹凤勤, 陆永跃
(1 华南农业大学 农学院, 广东 广州 510642; 2 浙江大学 农业与生物技术学院,浙江 杭州 310058; 3 海南大学 环境与植物保护学院, 海南 海口 570228)
橘小实蝇成虫复眼结构及感光机制
刘 欢1, 邓淑桢2, 赵晓峰1, 曹凤勤3, 陆永跃1
(1 华南农业大学 农学院, 广东 广州 510642; 2 浙江大学 农业与生物技术学院,浙江 杭州 310058; 3 海南大学 环境与植物保护学院, 海南 海口 570228)
【目的】研究橘小实蝇Bactroceradorsalis成虫复眼外部形态、内部显微结构和光感受机制。【方法】利用扫描电镜观察橘小实蝇成虫复眼外部形态,组织切片研究成虫复眼内部显微结构及在不同光环境下小眼结构的变化。【结果】橘小实蝇成虫复眼位于头部两侧,正面观呈半球形,表面光滑平坦;小眼间隙有感觉毛,单个小眼由角膜、晶锥、网膜细胞及其特化的视杆、基膜等构成,晶锥、视杆周围和色素细胞内均含有大量的色素颗粒。不同单色光处理后,小眼内的附属色素细胞色素颗粒沿小眼纵轴移动。白光、绿光和黄光处理后,附属色素细胞色素颗粒沿小眼纵轴均匀分布;紫光、蓝光和红光处理后,附属色素细胞色素颗粒主要集中在视网膜细胞远端和角膜的近端。【结论】橘小实蝇雌、雄成虫复眼在外形和内部结构上无差异,均属于并置像眼,屏蔽色素颗粒的移动是其复眼适应外界光环境变化的重要机制,该研究结果为筛选橘小实蝇成虫敏感光波并对其进行灯光诱杀提供了理论依据。
橘小实蝇; 复眼; 显微结构; 趋光性; 感光机制
橘小实蝇BactroceradorsalisHendel是一种世界性的重要果蔬害虫,可危害46个科250余种果树和蔬菜,雌、雄成虫存在多次交配行为,并具有强大的繁殖能力,常常爆发成灾,给果蔬业造成严重的经济损失[1]。因橘小实蝇幼虫潜居果实取食,老熟幼虫在土壤中化蛹,且田间种群对拟除虫菊酯、有机磷杀虫剂和阿维菌素等药剂的抗药性日益增强,致使传统的化学防治效果较差[2]。目前,利用甲基丁香酚诱杀雄虫是监测和防控橘小实蝇的主要技术之一[3]。
昆虫的复眼是其视觉系统的重要组成部分,对昆虫觅食、求偶、休眠、避害和决定行为方向等活动至关重要[4]。不同昆虫复眼的内部结构各不相同,但基本上都是由集光部分和感光部分组成,角膜和晶体为主要集光部分,起着传递和聚集光波的作用;视杆和视神经是主要感光部分。此外,在角膜和视杆周围还分布着大量的色素细胞,具有感受光波能量和产生神经冲动的作用[5]。
昆虫对光的趋性是视觉器官中的感光细胞接受光刺激后,对特定光源不可遏制的趋向行为,是昆虫长期对生态环境适应进化的结果[6-7]。桃小食心虫Carposinaniponensis成虫对波长350 nm的单色光趋性比较明显[8];亚洲小车蝗Oedaleusasiaticus和东亚飞蝗Locustamigratoriamanilensis对绿光、蓝光、紫光等短波段光波有强烈的趋光性[9];白背飞虱Sogatellafurcifera及其天敌黑肩绿盲蝽Cyrtorhinuslividipennis对蓝光均较为敏感[10];豆大蓟马Megalurothripsusitatus成虫对蓝色光的趋性显著高于其他波长光[11]。利用昆虫对不同光源的偏好特性可对害虫种群的种类与结构、数量与密度的变化、空间分布等进行监测与防治。
昆虫在不同光环境中进行觅食、飞行和躲避天敌等活动,都需要通过复眼对光波进行感应,因此研究复眼结构及其生理变化是了解昆虫趋光机制的理论基础。目前,国内外对一些重要的农业、林业害虫及天敌昆虫的复眼结构进行了大量的研究报道,如红火蚁Solenopsisinvicta[12]、大草蛉Chrysopapallens[13]、龟纹瓢虫Propyleajaponica[14]、拟天牛Xanthochroaluteipennis等[15]。关于橘小实蝇成虫复眼的显微结构及其光适应机制等鲜见相关研究报道。根据持续控制、安全有效的防治思路,本文以橘小实蝇成虫的趋光性作为切入点,利用扫描电镜和组织切片技术研究橘小实蝇复眼的外部形态、内部显微结构,及其在不同光环境下小眼结构的变化,试图阐明其复眼结构及感光机制,以期为灯光诱杀橘小实蝇的绿色防控措施提供依据。
1 材料与方法
1.1 材料
橘小实蝇为华南农业大学检疫与入侵害虫实验室使用人工饲料饲养的种群。使用配方改进的人工饲料饲养幼虫,用m(蔗糖)∶m(酵母粉)=1∶1的饲料饲养成虫。饲养室温度为27 ℃,相对湿度为(70±5)%,光周期为14h光∶10h暗。
LED灯:绿光(520~525 nm)、红光(620~625 nm)、黄光(590~595 nm)、蓝光(460~465 nm)、紫光(410~415 nm)、白光(6 000~6 500 K),电压为220 V,功率为3 W,光照度为1 500~2 000 lx(福州玛珂立电子有限公司);Nikon Eclipse CI正置光学显微镜、Nikon DS-U3成像系统[尼康映像仪器销售(中国)有限公司];日立H-3010扫描电子显微镜[日立(中国)有限公司]。
1.2 方法
1.2.1 橘小实蝇成虫复眼外部形态的观察 橘小实蝇成虫复眼外部形态观察参考那宇鹏等[16]的方法,将羽化20 d的橘小实蝇雌、雄成虫头部完整取下放入φ为70%乙醇溶液中,随后用超声波去除复眼表面污物,经φ为50%、70%、90%、95%和100%的乙醇溶液梯度脱水,干燥后用导电胶固定样品,SBC-11型离子溅射镀膜仪内喷金,置于扫描电子显微镜下观察并拍照。
1.2.2 橘小实蝇成虫复眼显微结构的观察 橘小实蝇成虫复眼显微结构观察参考李琼艳等[5]和张胜杰等[17]方法,将羽化20 d的雌、雄成虫在白光、绿光、黄光、紫光、蓝光、红光下处理3 h,快速将其头部切入φ为4%多聚甲醛溶液中固定24 h以上,于脱水机内依次φ为75% 乙醇溶液4 h、φ为85%乙醇溶液2 h、φ为90%乙醇溶液2 h、φ为95%乙醇溶液1 h、无水乙醇 30 min进行梯度脱水,然后浸蜡并于包埋机内进行包埋,将修整好的蜡块置于石蜡切片机上切片,片厚为4 μm,HE染色晾干后用中性树胶封片,光学显微镜下观察并拍照。
2 结果与分析
2.1 橘小实蝇成虫复眼的外部形态
橘小实蝇雌、雄成虫头部及复眼外部形态见图1。雌、雄成虫复眼形状、大小无明显差异,均位于头部两侧,正面观呈半球形,有光泽,扫描电镜下观察可见复眼表面光滑平坦(图1A、1F);昆虫的复眼由多个小眼组成,复眼中心区域小眼基本属于典型的六边形结构,边缘区域的小眼为规则的四边形,各小眼面微凸,排列整齐、紧密,相互拼合在一起,似蜂窝状,小眼间隙附有感觉毛,多垂直于复眼表面,少数端部呈弯曲状(图1E、1J)。

A:雄虫头部实物图;B:雄虫头部全貌;C:雄虫单个复眼全貌;D:雄虫复眼局部放大;E:雄虫复眼小眼紧密排列(箭头示感觉毛);F:雌虫头部实物图;G:雌虫头部全貌;H:雌虫单个复眼全貌;I:雌虫复眼局部放大;J:雌虫复眼小眼紧密排列(箭头示感觉毛)。
图1 橘小实蝇雌、雄成虫复眼的外部形态
Fig.1 The external morphology of compound eyes ofBactroceradorsalisfemale and male adults
2.2 橘小实蝇成虫复眼的内部显微结构
橘小实蝇成虫复眼的显微结构组织切片见图2,由图2可知,橘小实蝇成虫复眼由大量的小眼组成,每个小眼由角膜、晶锥、视网膜细胞、视杆、屏蔽色素细胞和基膜等部分构成。小眼的最外层为角膜,由角膜细胞向外分泌而成,表面光滑,纵切面为外缘向外突出、内部稍凹的凸透镜透明结构(图2C,2E);角膜下方是晶锥细胞分泌形成的透明并具有折光性的晶锥,与角膜组成屈光系统,末端与视杆中心的视杆相连(图2C,2H),其纵切面呈不规则的倒锥状,横切面呈不规则多边形(图2F,2G),周围分布色素颗粒;橘小实蝇成虫复眼有主色素细胞和附属色素细胞2种不同类型的屏蔽色素细胞,主色素细胞包裹晶锥外围,其色素颗粒呈黄色,而附属色素细胞包裹整个小眼,其色素颗粒呈红色(图2A~F);视杆由视网膜细胞特化而成,呈离散型分布于晶锥和基膜之间(图2H),是光感受器的主要感光部位,与晶锥直接相接触,一直延伸到小眼的近基膜处;基膜位于小眼最底层,是一层结缔组织,呈栅栏状,是连接小眼和视神经的部分,起到机械性支撑小眼的作用,同时能将透入的光波再次反射到视杆上,增加视神经的感受性,基膜底部分布有大量的屏蔽色素颗粒。
2.3 不同光环境下橘小实蝇成虫复眼的显微结构
在不同波长光处理后,主色素细胞屏蔽色素颗粒无明显变化,附属色素细胞屏蔽色素颗粒随着不同波长光的处理而发生纵向移动(图3)。白光、绿光、黄光处理后,附属色素细胞屏蔽色素颗粒遍布小眼纵轴,均匀地分布在视杆两侧上下(图3A~3C);紫光、蓝光处理条件下,在视网膜细胞远端和靠近角膜的近端屏蔽色素颗粒较多,视杆中段屏蔽色素颗粒较少(图3D~3E);红光处理后,屏蔽色素颗粒大量集中在视网膜细胞远端和靠近角膜的近端(图3F)。不同光环境下,橘小实蝇雌、雄成虫复眼的显微结构未见明显差异。
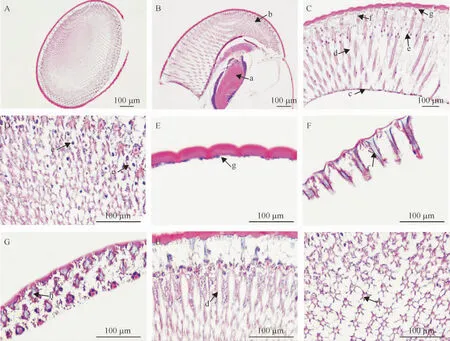
A:复眼横切;B:复眼纵切(箭头示a:视叶;b:复眼);C:复眼结构(箭头示c:基膜;d:视杆;e:色素细胞;f:晶锥;g:角膜);D:色素颗粒(箭头示e:色素细胞);E:角膜纵切(箭头示g:角膜);F:晶锥纵切(箭头示f:晶锥);G:晶锥横切(箭头示h:晶锥);H:视杆远心端纵切(箭头示d:视杆);I:视杆中段横切(箭头示i:视杆)。
图2 橘小实蝇成虫复眼显微结构组织切片
Fig.2 The microstructures of compound eyes ofBactroceradorsalisadults revealed by paraffin section
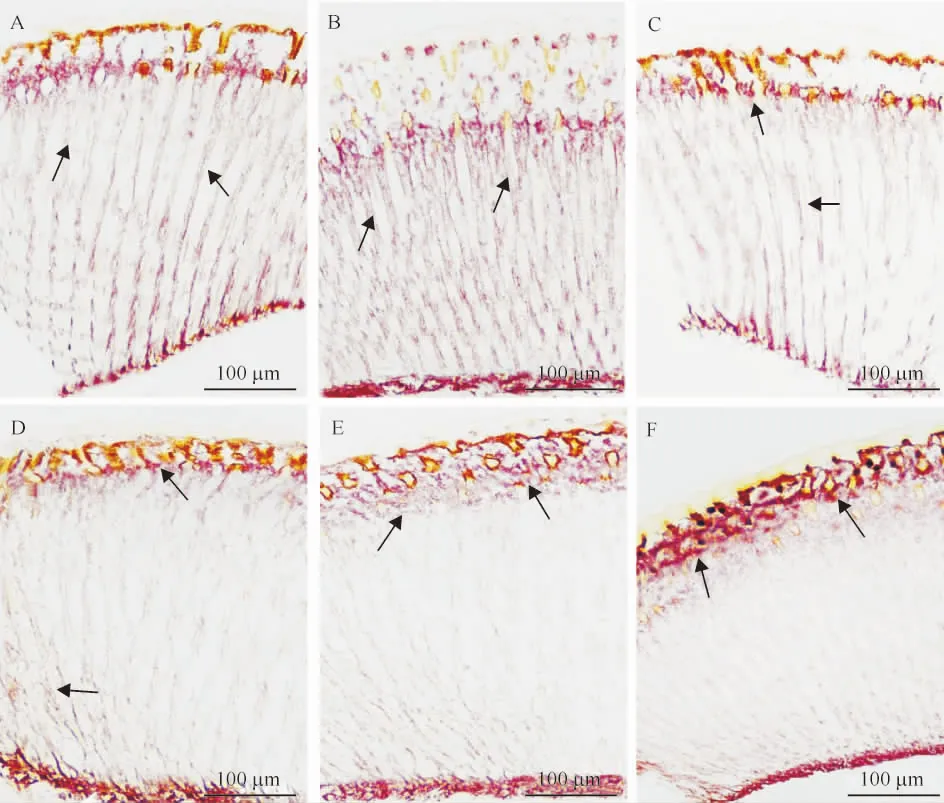
A:白光处理,B:绿光处理,C:黄光处理,D:紫光处理,E:蓝光处理,F:红光处理;箭头示色素颗粒。
Fig.3 Longitudinal sections of several ommatidia of compound eyes ofBactroceradorsalisadults treated by lights of different wavelength (unstained)
3 讨论与结论
昆虫复眼具有辨识图案或识别形状的能力,尤其是对运动的物体有较高的辨别力,对善于飞翔的双翅目昆虫发现、识别、跟踪、捕获对它们有意义的目标至关重要[18]。昆虫复眼结构的研究对昆虫的分类鉴定、防治及行为学研究具有重要意义,也是探索昆虫视觉行为与外界环境相互关系的理论基础[18-19]。本研究结果表明,橘小实蝇雌、雄成虫复眼在外部形态、小眼数量、大小及排列方式方面均无明显差异。昆虫复眼的小眼数量往往是影响复眼视野、视觉敏锐度的重要因素[17],橘小实蝇成虫复眼占据除额沿外大部分头部,单个复眼约有3 800个小眼[20],使其具有广阔的视野,有利于提高成虫寻找寄主植物、交配和躲避天敌等生命行为的成功率。
昆虫复眼表面一般着生感觉毛、乳状突起颗粒和钟形感觉器等辅助结构,鳞翅目昆虫家蚕Bombyxmori和美国白蛾Hyphantriacunea成虫复眼的角膜外表面有一些非常微小的乳头状突起,具有减少光线的反射并增加感受能力的作用[5];直翅目昆虫双斑蟋Gryllusbimaculatus复眼表面具有少而短的机械感觉毛和钟形感觉器来感知外界机械感受力[16]。橘小实蝇复眼表面光滑不具有乳状突起颗粒和钟形感觉器,但小眼间空隙随机分布着白色感觉毛,这种特征和擅长飞行类昆虫的复眼结构相似,据报道,多种蝶类复眼表面都具有长的刺毛,在飞行时可以感受风速的变化[21],复眼感觉毛在蛆症异蚤蝇Megaseliascalaris飞行过程中起机械感觉作用[22],蜜蜂复眼表面的感觉毛可以帮助其精确测量取食场所与太阳方向之间的角度[23]。
橘小实蝇成虫每个小眼都是一个独立的视觉单位,主要包括角膜、晶锥、视网膜细胞、视杆、色素细胞和基膜等结构。昆虫复眼的结构特征与其生物习性相关,日间活动昆虫复眼的晶锥与视杆直接相连,两者之间没有无色素颗粒的透明区域,为并置像眼;夜间活动昆虫复眼的小眼延长,视杆远离晶锥,两者之间存在无色素颗粒的透明区域,为重叠像眼[18,24]。根据橘小实蝇成虫复眼显微结构及生活习性,推测其复眼属于并置像眼,与螺旋粉虱Aleurodicusdisperses[18]、异色瓢虫Harmoniaaxyridis[25]、双斑蟋成虫[16]的复眼结构相似。
昆虫的复眼对不同波长的光都有感知能力,能够感受到偏振光且具有色觉[26]。但不同种类的昆虫对不同波长的单色光敏感性差异较大,在不同光环境下,昆虫复眼通过其色素细胞内屏蔽色素颗粒的移动调整复眼状态,以适应外界光环境的变化[6]。在白光、绿光和黄光条件下,橘小实蝇成虫复眼小眼内附属色素细胞内的屏蔽色素颗粒较均匀地分布于视杆两侧上下,维持正常的视觉功能;在蓝光、紫光和红光条件下,屏蔽色素颗粒向小眼远端移动,在晶锥处聚集,提高复眼的绝对敏感度,与螺旋粉虱[18]、龟纹瓢虫[14]、异色瓢虫[25]和黑带食蚜蝇Episyrphusbalteatus[17]等成虫复眼的光适应机制相同。
昆虫复眼视网膜细胞是其感光的重要场所,当感受到外界光刺激时,其膜电位通常会发生改变。利用视网膜细胞电位技术可以测定光刺激后视网膜细胞电位变化的规律,从而筛选出昆虫的敏感光谱[6]。本研究发现在绿光和黄光处理时橘小实蝇成虫复眼的小眼显微结构变化与白光环境下相似,初步推测成虫对绿光和黄光具有较强的敏感性。已有研究表明,橘小实蝇成虫具有较强的趋光性和颜色偏好性,光刺激是其成虫飞翔活动的基本条件[27],田间调查发现橘小实蝇雌成虫偏好在接近成熟的黄色果实上产卵[28],室内选择性试验发现橘小实蝇成虫对绿色的食物具有较强的趋性[29]。关于橘小实蝇成虫复眼对不同波长光刺激的视网膜细胞电位变化及成虫趋光反应行为等还需进一步研究。
[1] SHEN G M, WANG X N, DOU W, et al. Biochemical and molecular characterization of acetylcholinesterase in four field populations ofBactroceradorsalis(Hendel) (Diptera:Tephritidae)[J]. Pest Manag Sci, 2012, 68(12): 1553-1563.
[2] JIN T, ZENG L, LIANG G W, et al. Insecticide resistance of the oriental fruit fly,Bactroceradorsalis(Hendel) (Diptera: Tephritidae), in mainland China[J]. Pest Manag Sci, 2011, 67(3): 370-376.
[3] PAGADALA D K J, CHRITINE M W, JOHN C, et al. Isolation and identification of host cues from Mango,Mangiferaindica, that attract gravid female oriental fruit fly,Bactroceradorsalis[J]. J Chem Ecol, 2012, 38(4): 361-369.
[4] STALLEICKEN J, LABHART T, MOURITSEN H. Physiological characterization of the compound eye in monarch butterflies with focus on the dorsal rim area[J]. J Comp Physiol, 2006, 192(3): 321-331.
[5] 李琼艳,刘春,夏庆友,等. 家蚕复眼突变系光泽眼(lu)和光泽小眼(ve)的复眼形态观察[J]. 昆虫学报,2013,56(4):350-357.
[6] 于海利. 梨小食心虫GrapholitamolestaBusck的趋光性及绿光对其生物学特性的影响[D]. 杨凌:西北农林科技大学, 2011.
[7] 蒋月丽,武予清,李彤,等. 铜绿丽金龟对不同光谱的行为反应[J]. 昆虫学报, 2015, 58(10):1146-1150.
[8] 侯无危,马幼飞,高慰曾,等. 桃小食心虫蛾的趋光性[J]. 昆虫学报, 1994, 37(2):165-170.
[9] 牛虎力,周强. 光电诱导作用下蝗虫的光谱和光强反应行为研究[J]. 现代生物医学进展, 2009, 9(3):466-469.
[10]邵英,程建军,刘芳. 白背飞虱及其天敌黑肩绿盲蝽的趋光性研究[J]. 应用昆虫学报, 2013, 50(3):700-705.
[11]唐良德,韩云,吴建辉,等. 豆大蓟马室内对不同颜色及光波的趋性反应[J]. 植物保护, 2015, 41(3):169-172.
[12]FAN F, ZHAO C, REN H, et al. External morphology and microstructure of the compound eye of fire ants,SolenopsisinvictaBuren[J]. Front Agric China, 2011, 5(4):570-575.
[13]张海强,朱楠,魏国树,等. 大草蛉成虫复眼的外部形态及其显微结构[J]. 昆虫学报, 2007, 50(5):454-460.
[14]闫海燕,魏国树,闫海霞,等. 龟纹瓢虫成虫的复眼形态及其显微结构[J]. 昆虫知识, 2006, 43(3):344-348.
[15]MISHRA M, MEYER-ROCHOW V B. Eye ultrastructure in the pollen-feeding beetle,Xanthochroaluteipennis(Coleoptera: Cucujiformia: Oedemeridae)[J]. J Electron Microsc, 2006, 55(6):289-300.
[16]那宇鹏,冷雪,那杰. 双斑蟋复眼和视叶的显微结构[J]. 应用昆虫学报, 2015, 52(6):1482-1490.
[17]张胜杰,范凡,魏国树,等. 黑带食蚜蝇EpisyphusbalteatusDe Geer的复眼结构及其调光机制[J]. 应用昆虫学报, 2015, 52(3):750-758.
[18]时艳华,吴伟坚,符悦冠. 螺旋粉虱成虫的复眼形态及其内部结构[J]. 环境昆虫学报, 2010, 32(1):41-46.
[19]张晓俞,马俊宁,陈辉. 华山松大小蠹成虫复眼的外部形态及显微结构[J]. 西北农林科技大学学报(自然科学版), 2015, 43(9):109-114.
[20]XUE H, ZHENG L X, WU W J. Morphometry of compound eyes of threeBactrocera(Diptera: Tephritidae) species[J]. Fla Entomol, 2015, 98(2): 807-809.
[21]陈伟之,杨思威,李素梅,等. 蝶类复眼小眼面表面超微结构的比较研究[J]. 昆虫学报, 2002, 45(1):35-40.
[22]LAU T F S, OHBA N, ARIKAWA K, et al. Sexual dimorphism in the compound eye ofRhagophtalmusohbai(Coleoptera: Rhagophthalmidae): II: Physiology and function of the eye of the male[J]. J Asia-Pac Entomol, 2007, 10(1): 27-31.
[23]NEESE V. Zur Funktion der Augenborsten bei der Honigbiene[J]. Z Vergl Physiol, 1965, 49(6): 543-585.
[24]MEYER-ROCHOW V B, GáL J. Dimensional limits for arthropod eyes with superposition optics[J]. Vision Res, 2004, 44(19): 2213-2223.
[25]吴春娟,陈洁,范凡,等. 异色瓢虫显现变种复眼的形态、显微结构及其光暗条件下的适应性变化[J]. 昆虫学报, 2011, 54(11):1274-1280.
[26]冷雪,那杰. 昆虫复眼的结构和功能[J]. 沈阳师范大学学报(自然科学版), 2009, 27(2):241-244.
[27]刘建宏,叶辉. 光照、温度和湿度对桔小实蝇飞翔活动的影响[J]. 昆虫知识, 2006, 43(2):211-214.
[28]黄阿凤,林良方. 漳州地区柑橘小实蝇危害状况和防治对策[J]. 福建热作科技, 2004, 29(1):33-34.
[29]梁丹辉,陈冉,周琼. 桔小实蝇成虫对不同颜色和气味物质的趋性反应[J]. 生命科学研究, 2012, 20(1):36-39.
【责任编辑 霍 欢】
Structure and photoreception mechanism of the compound eye of Bactrocera dorsalis Hendel
LIU Huan1, DENG Shuzhen2, ZHAO Xiaofeng1, CAO Fengqin3, LU Yongyue1
(1 College of Agriculture, South China Agricultural University, Guangzhou 510642, China;
2 College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310058,China;
3 College of Environment and Plant Protection, Hainan University, Haikou 570228, China)
【Objective】 To study the external morphology, internal microstructure and photoreception mechanism of compound eye of the oriental fruit fly (Bactroceradorsalis).【Method】The external morphology of compound eye ofB.dorsalisadult was observed using a scanning electron microscope. The microstructure of compound eye and the structural change of ommatidia under different monochromatic lights were observed using histological biopsy.【Result】The compound eyes were hemispheric, smooth, and located on the lateral upside of head. The gaps among ommatidia were covered with interommatidial hairs. Each ommatidium was comprised of a cornea, a crystalline cone, retinula cells, a rhabdom and a basement membrane. There were abundant pigment granules in the retinular pigment cells, and in the surroundings of the crystalline cone and rhabdom. The pigment granules in accessory pigment cells moved longitudinally under different monochromatic lights. They were distributed uniformly along the longitudinal axis in the ommatidium under white, green and yellow light, while mainly clustered in the distal of retinular cells and proximal of cornea under violet, blue and red light.【Conclusion】There are no difference in external morphology and internal microstructure of compound eyes between male and female ofB.dorsalis, and the compound eyes of both sexes are apposition eyes. Their compound eye adapts to environmental light changes by the mechanism of pigment granules moving longitudinally. This research can provide a theoretical basis for screening sensitive lights ofB.dorsalisadult for light trapping.
Bactroceradorsalis;compound eye;microstructure;phototaxis;photoreception mechanism
2016- 06- 03优先出版时间:2017-01-10
刘 欢(1987—),男,博士研究生,E-mail:liuhuan@stu.scau.edu.cn;通信作者:陆永跃(1972—),男,教授,博士,E-mail:luyongyue@scau.edu.cn
国家重点研发计划项目(2016TFC1201200);广东省高等学校高层次人才项目(粤财教[2013]246号)
Q968
A
1001- 411X(2017)02- 0075- 06
优先出版网址:http://www.cnki.net/kcms/detail/44.1110.s.20170110.1423.026.html
刘 欢, 邓淑桢, 赵晓峰, 等.橘小实蝇成虫复眼结构及感光机制[J].华南农业大学学报,2017,38(2):75- 80.
