信号分子对汤姆青霉PT95菌株菌核分化的影响
2017-01-05赵文婧韩建荣
赵文婧,韩建荣
(1.太原师范学院生物系,山西太原030031;2.山西大学生命科学学院,山西太原030006)
信号分子对汤姆青霉PT95菌株菌核分化的影响
赵文婧1,韩建荣2
(1.太原师范学院生物系,山西太原030031;2.山西大学生命科学学院,山西太原030006)
从菌核分化入手,研究外源信号分子对汤姆青霉PT95菌株菌核分化的影响。使用MEA和PDA这2种不同的培养基培养汤姆青霉PT95菌株,并在2种不同的培养基上分别添加0,0.02,0.04,0.06 mmol/mL的cAMP;通过记录菌核成熟的时间,测量菌落直径、菌核生物量和类胡萝卜素含量,以分析菌株生长发育情况。结果表明,在加入的cAMP浓度达到0.02 mmol/mL时菌核的生长发育状态最佳;之后随着cAMP浓度的增加,其生长状况逐渐下降;当cAMP浓度达到0.06 mmol/mL时,其生长开始受到抑制,表明外界适当供给信号分子cAMP可促进汤姆青霉PT95菌株的生长,但cAMP浓度过高则会抑制汤姆青霉菌株的生长发育。
cAMP;汤姆青霉;菌核;类胡萝卜素
菌核是由菌丝绞缠聚集而形成的坚硬的休眠结构,真菌学家和植物病理学家对真菌菌核发育和结构的研究主要集中在子囊菌和担子菌[1-2]。据《中国真菌志》第三十五卷[3]记载,从我国分离到的8种能产生菌核的青霉中包括汤姆青霉(Penicillium thomii)。从土壤中分离到的一株经鉴定属于Penicillium thomii的汤姆青霉PT95菌株,能在普通的固体培养基(即琼脂平板)上形成大量坚硬的砂粒状的微菌核[4],并在菌核的脂质小球中积累大量的以β-胡萝卜素为主要成分的类胡萝卜素。在进行青霉的形态观测时,主要包括3个方面:菌落的生长速率,即在一定时间内一定的培养基上的菌落直径;菌落的特征;显微特征[5]。
有研究发现,一些外源信号分子可延缓果蔬衰老[6]。生物体病变衰老时体内的自由基会增加,类胡萝卜素能够抑制自由基的形成,延缓衰老和预防癌症。近年来,利用微生物生产类胡萝卜素也已经有了商业化应用[7]。β-胡萝卜素可以提高种鸡产蛋率和孵化率[8],是一种有效的过氧化自由基的捕获剂。当活性氧(Reactive Oxygen Species,ROS)在体内积累浓度过高时会导致细胞损伤,而β-胡萝卜素含多个共轭双键,能淬灭单线态氧(1O2),以避免1O2对脂类双键或共轭双键的攻击;清除自由基来阻碍或终止反应链的进行[9],使得其具有抗氧化损伤的能力。
菌丝中储存的营养成分酶解后可以为菌核发育功能提供能量,形成菌核的大小、数量与培养基中的营养构成有关[10]。葡萄孢菌(Botrytis cinerea)野生型一般不产生菌核,但其突变株cAMP的水平显著降低,可产生大量的菌核;若加入cAMP,会抑制菌核的形成,表现出野生型的性状。说明cAMP对B.cinerea菌核发育起着负调控作用。cAMP参与菌核的发育在不同的种群中有差异,PT95菌株在各种不同的培养基上均表现出理想的选择培养特性[11],而汤姆青霉PT95在菌核中可以积累大量的类胡萝卜素[12]。故本试验采用汤姆青霉PT95菌株进行研究,探索cAMP浓度对其菌核分化的影响。
通过查阅相关文库发现,关于PT95菌株菌核分化与类胡萝卜素在高氧胁迫光照条件下的产率[13]及生物诱导方面的理论研究[14]已有记录,但没有汤姆青霉PT95菌株对外源信号分子细胞响应的研究报道,所以,本研究有一定的创新性,不仅可对汤姆青霉PT95菌株菌核分化的信号传递机制有更多认识,也能对汤姆青霉PT95菌株的菌核分化提供更完整的依据,为产菌核真菌的研究奠定一定的理论基础。
1 材料和方法
1.1 材料
1.1.1 菌株Penicillium thomii PT95菌株,分离自山西汾阳的混交林土壤,保存在查氏(CA)斜面上。
1.1.2 培养基麦芽汁培养基(MEA):麦芽膏20 g,蛋白胨1.0 g,葡萄糖20 g,琼脂20 g,蒸馏水1 L。
马铃薯葡萄糖琼脂培养基(PDA):20%马铃薯煮汁(称取马铃薯200 g,加水1 000 mL,煮沸20 min后,过滤。在滤汁中补足水分到1 000 mL,即成20%马铃薯煮汁)1 000 mL,葡萄糖20 g,琼脂18 g。
1.1.3 主要仪器设备灭菌锅,培养箱,超净台,电子天平,Canon相机,电磁炉,高压蒸汽灭菌锅。
1.2 方法
1.2.1 配制培养基MEA培养基适合PT95菌株大量产生菌核,而在PDA培养基上其类胡萝卜素产率最高[15]。按照配方配制好培养基后,分装到若干个三角瓶中,封好放入高压灭菌锅中,在121℃条件下灭菌20 min。
1.2.2 不同浓度cAMP的设置在MEA和PDA培养基中添加cAMP,使得培养基中cAMP的浓度分别达到0.02,0.04,0.06 mmol/mL;同时以没有添加cAMP的培养基作为对照。每个9 cm培养皿中倒入25 mL培养基。
1.2.3 接种MEA和PDA培养基按浓度梯度各分为4组,分别记为A,B,C,D,用移液枪分别添加0,0.02,0.04,0.06 mmol/mL的cAMP试剂5 μL,每组设置3个重复。在超净工作台上,用接种针挑取汤姆青霉PT95菌株的单菌核,以三点式接种方法接入培养皿中,分别置于25℃培养箱中黑暗培养。
1.2.4 菌落形态及特征观察观察不同培养基上PT95菌株的菌落特征,记录渗出液出现和消失的时间,在第7天及菌核成熟后用Canon数码相机自然光下照相。待渗出液渐渐消失,即菌核成熟之时,收集PT95菌株在不同培养基上的成熟菌核,用标尺测量2种培养基上的2株菌株成熟菌落的直径。
1.3 测定项目及方法
1.3.1 菌核生物量测定用自来水将青霉PT95菌株的成熟菌核从平板上冲洗下来,收集到平皿中,烘干称质量。
1.3.2 类胡萝卜素的提取及含量测定根据韩建荣等[4]的方法提取青霉PT95菌株成熟菌核中的类胡萝卜素,然后按王业勤等[16]的方法计算所提取菌核中的类胡萝卜素含量。即将干菌核置于研钵中进行研磨,再加入5 mL丙酮、10 mL氯仿进行提取,4 000 r/min离心10 min。取上清液加10 mL蒸馏水,振荡、静置分层,取下层约5 mL收入容量瓶,用丙酮定容至10 mL。在475 nm下,以丙酮溶液作为空白对照,将提取好的液体用分光光度计测定吸光度值。
类胡萝卜素含量(μg/g)=(吸光度值×总体积(mL)×稀释倍数×10)/菌核干质量×2 500。
2 结果与分析
2.1 不同浓度cAMP对菌落特征和菌核分化的影响
在MEA和PDA培养基上PT95菌株的菌落表面几乎由菌核覆盖,只是在菌落边缘部分有非常少量的分生孢子。在MEA培养基上,汤姆青霉PT95菌株在第4天出现了渗出液(大量菌丝集结形成菌核,生长发育使菌丝相互挤压时,菌丝细胞液渗出,即为渗出液[17]);第6天渗出液渐渐消失,菌核出现,菌丝交缠聚集;第11天菌核成熟。在cAMP浓度为0.02 mmol/mL时,渗出液出现、消失及菌核成熟的时间较对照都提前了1~2 d,说明添加cAMP后对菌株生长有促进作用;但随着cAMP浓度的增加,渗出液出现、消失及菌核成熟时间渐渐推迟;当cAMP浓度为0.06 mmol/mL时,菌核渗出液出现及菌核成熟的时间都较对照推迟了1 d,说明添加较高浓度的cAMP会抑制菌株的生长。
由表1可知,在PDA培养基上,汤姆青霉PT95菌株在第5天出现了渗出液;第7天渗出液渐渐消失,菌核出现,菌丝交缠聚集;第10天菌核成熟。在cAMP浓度为0.02 mmol/mL时,渗出液出现、消失及菌核成熟的时间都较对照有所提前;当cAMP浓度为0.04 mmol/mL时,菌核生长状况与对照基本相同;当cAMP浓度为0.06 mmol/mL时,菌核渗出液出现及菌核成熟的时间都较对照推迟。
综上所述,PT95菌株在MEA培养基上比在PDA培养基上渗出液和菌核出现的时间提早1 d;但菌核成熟的时间,MEA培养基比PDA培养基要推迟1 d。
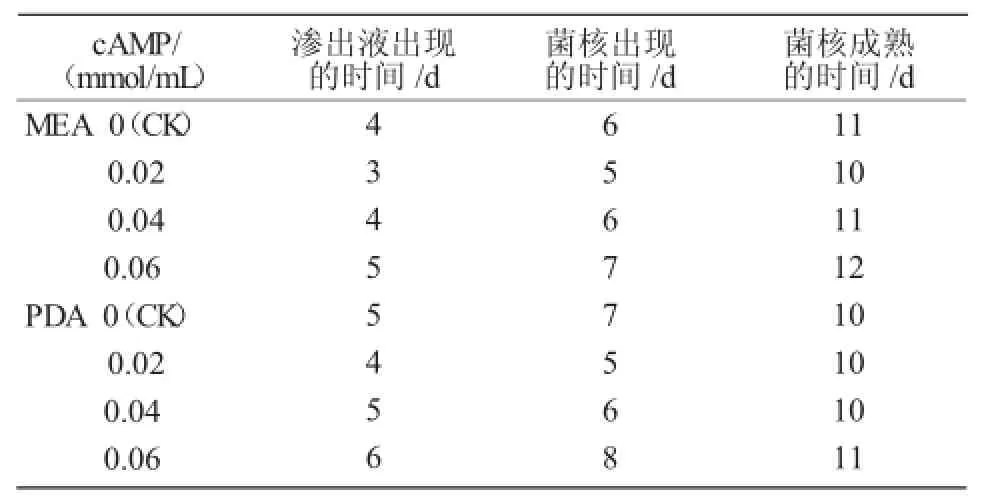
表1 不同浓度cAMP对PT95菌核分化的影响
2.2 不同浓度cAMP对菌核形态和菌核生物量的影响
通过测量,汤姆青霉PT95菌株菌核形状以圆形为主(图1)。

由表2可知,PT95菌株在MEA培养基比PDA培养基上的菌落直径要大一些,且随着cAMP浓度的增加,MEA和PDA培养基上的菌落直径均先增后减,当cAMP浓度为0.02 mmol/mL时,MEA和PDA培养基上的菌核生长均最好;之后菌核生长不再受促进,当cAMP浓度为0.06 mmol/mL时,菌核生长受到抑制。
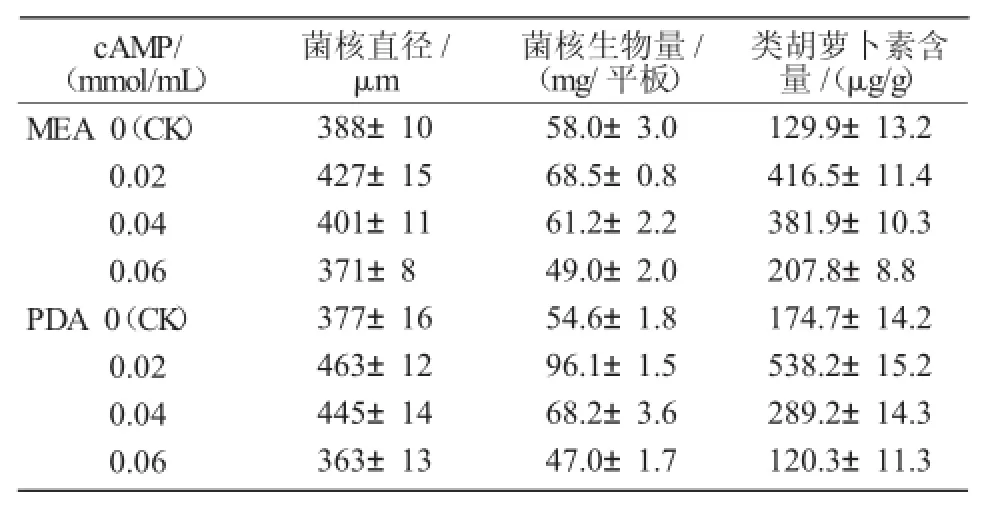
表2 不同浓度cAMP对PT95菌核分化的影响
从表2还可以看出,PT95菌株在MEA培养基上的菌核生物量比在PDA培养基上稍高一些,达到58.0 mg/平板;随着cAMP浓度的变化,菌核的生物量也在变化。其中,在MEA培养基上,当cAMP浓度为0.02 mmol/mL时,菌核生物量达到最高值,为68.5 mg/平板,是对照的1.18倍;然后逐渐降低;当cAMP浓度为0.06 mmol/mL时降至最低值,是对照组的84%。在PDA培养基上,菌核生物量相对较低一些,为54.6 mg/平板;当cAMP浓度为0.02 mmol/mL时达到最高值,为96.1 mg/平板,是对照的1.76倍;当cAMP浓度为0.06 mmol/mL时降至最低值,低于对照组,是对照组的86%。
综上所述,随着外源信号分子cAMP浓度的增加,PT95菌株菌核直径和菌核的生物量均呈先增大后减小的趋势。
2.3 不同浓度cAMP培养基对菌核类胡萝卜素含量的影响
由表2可知,在MEA培养基上,PT95菌株的类胡萝卜素含量达到129.9 μg/g。在添加不同浓度的cAMP时,PT95菌株的类胡萝卜素含量有所不同,当cAMP浓度为0.02 mmol/mL时,类胡萝卜素含量达到最高值,为416.5 μg/g,是对照的3.2倍;然后随着cAMP浓度的增加,类胡萝卜素含量逐渐下降;当cAMP浓度为0.06 mmol/mL时,其含量降至最低值,为207.8 μg/g,但仍然高于对照,是对照的1.6倍。
在PDA培养基上,PT95菌株的类胡萝卜素含量达到174.7 μg/g。当添加cAMP的浓度为0.02 mmol/mL时,类胡萝卜素含量达到最高值,为538.2 μg/g,是对照组的3.1倍;随着cAMP浓度的增加,类胡萝卜含量逐渐下降;当cAMP浓度为0.06 mmol/mL时,其含量降至最低值,为120.3 μg/g,为对照组的70%。
综上所述,随着外源信号分子浓度的提高,类胡萝卜素含量呈先升高后降低的趋势;当cAMP浓度为0.02 mmol/mL时,类胡萝卜素含量最高,明显高于对照,表明cAMP对提高类胡萝卜素含量有促进作用;当cAMP浓度继续增加时,含量有所下降,cAMP浓度为0.06 mmol/mL时,类胡萝卜素含量低于对照,说明菌核生长受到抑制。
3 结论与讨论
本试验结果表明,MEA培养基比PDA培养基培养菌落时渗出液出现时间早,菌核也较早出现;但PT95菌株在PDA培养基上成熟的时间要比在MEA培养基上更早一些。从培养效果上看,MEA培养基比PDA培养基更适合于使汤姆青霉PT95菌株产生大量的菌核。
有研究表明,使细胞内cAMP含量升高的因素会降低细胞生长速度,抑制细胞的增殖,使失去接触抑制的细胞恢复控制性生长[18]。对巨大芽孢杆菌而言,外源cAMP几乎不影响其生长;而对E.coli AS 1.797菌株以葡萄糖作碳源来讲,外源cAMP的存在对生长有抑制作用,说明外源cAMP可能对不同菌株有着不同的作用[19]。
本研究结果表明,cAMP的浓度对汤姆青霉菌株的分化有一定的影响,在培养基上,随着cAMP浓度的提高,2株菌株出现渗出液的时间、出现菌核的时间均提前1~2 d,而菌核成熟的时间出现不同程度的推迟现象。外源信号分子浓度对汤姆青霉PT95菌株的菌核形态也有一定的影响,随着cAMP浓度的提高,菌株的菌落直径先增加后减少。这种变化有可能是对cAMP浓度的一种适应性反应,较低浓度的cAMP会适当促进菌核生长,但是浓度增加到一定程度则会对菌核生长产生抑制作用。
在本试验条件下,汤姆青霉PT95菌株的菌核生物量以及菌核中类胡萝卜素的含量与外源信号分子浓度呈一定的正相关,随着cAMP浓度的增加类胡萝卜含量先升高后降低。可能是由于汤姆青霉PT95和Q1菌株的机体中其他抗氧化系统起到了一定的作用,并不需要合成太多的类胡萝卜素来清除ROS,因此,类胡萝卜素含量有少许下降。
一般来说,当真菌消耗完碳源的时候,也就同时失去了对其抗氧化保护机制的有效管理能力,所以,产菌核真菌通过分化形成菌核以备长期生存。本试验结果表明,在外界适当供给cAMP条件下,汤姆青霉PT95菌株生长效果受到促进:一方面在菌核中积累类胡萝卜素这种抗氧化剂来保护机体,另一方面也能形成大量的菌核或者增加菌核的大小来帮助其度过不良环境。但是如果供给cAMP的浓度过高,PT95菌株的菌核生物量和类胡萝卜素含量反而会低于正常情况,其生长受到抑制。
今后在进行汤姆青霉PT95菌株的培养时,为了增强培养效果,可以使用MEA培养基添加适量浓度的cAMP,其中,0.02 mmol/mL的浓度下菌落生长状况最佳,而后随着其浓度增加而渐趋下降;当cAMP浓度达到0.06 mmol/mL时,菌株生长受到抑制。
[1]Luttrell E S.Host-parasite relationships and development of the ergot sclerotium in Claviceps purpurea[J].Canadian Journal of Botany,1980,58:942-958.
[2]Chet I,Henis Y.Sclerotial morphogenesis in fungi[J].Annual Review of Phytopathology,1975,13:169-192.
[3]孔华忠.中国真菌志(第三十五卷,青霉属及其相关有性型属)[M].北京:科学出版社,2007.
[4]韩建荣,王肖娟,原香娥.青霉PT95菌株菌核内产生类胡萝卜素的研究[J].微生物学通报,1998,25(6):319-321.
[5]Pitt J I.An appraisal of identification methods for Penicillium species:novel taxonomic criteria based on temperature and water relations[J].Mycologia,1973,65:1134-1157.
[6]李姗姗,王明力,吴映梅.外源信号分子在果蔬保鲜中的应用[J].贵州农业科学,2013,41(8):160-162.
[7]姜文侯,单志萍,孟妤.β-胡萝卜素的应用、市场和天然型产品的发酵法生产[J].食品与发酵工业,1994,20(3):65-71.
[8]范永仙,许尧兴.微生物生产类胡萝卜素的研究进展[J].食品与发酵工业,2003,29(7):69-74.
[9]周丽,梁新乐,励建荣.类胡萝卜素抗氧化作用研究进展[J].食品研究与开发,2003,24(2):21-23.
[10]Willetts H J,Wong A L.Ontogenetic diversity of sclerotia of Sclerotinia sclerotiorum and related species[J].Transactions of the British Mycological Society,1971,57:515-524.
[11]崔丽霞.青霉PT95菌株的鉴定和培养条件的研究[D].太原:山西大学,2004.
[12]韩建荣,李智伟,王宇,等.一株产菌核青霉的生物学特性研究[J].山西大学学报,2009,3(4):634-639.
[13]赵文婧.外源β-胡萝卜素、光照对青霉PT95菌株菌核分化和类胡萝卜素产率的影响[J].微生物学报,2005,45(2):279-282.
[14]韩建荣,高宇英,赵文婧.几种诱导子对青霉PT95菌株固态发酵产生类胡萝卜素的影响[J].应用与环境生物学报,2005,11(2):208-210.
[15]赵文婧,张泽清,龙丹丹,等.汤姆青霉PT95和Q1菌株产菌核培养基的筛选[J].山西大学学报,2013,36(4):622-627.
[16]王业勤,李勤生.天然类胡萝卜素研究进展、生产、应用[M].北京:中国医药科技出版社,1997.
[17]戚敬.青霉PT95菌株渗出液的产生以及菌核发育和类胡萝卜素代谢的关系[D].太原:山西大学,2006.
[18]高士争,张曦,程美玲.环腺苷酸的生物学作用与动物营养代谢调控[J].中国畜牧兽医,2003,30(6):21-23.
[19]颜日祥,段康明.环化腺苷酸对细菌生长的影响[J].微生物学报,1989,29(1):20-22.
Effects of Signal Molecule on Sclerotial Differentiation of Penicillium thomii PT95
ZHAO Wen-jing1,HAN Jian-rong2
(1.Department of Biology,Taiyuan Normal University,Taiyuan 030031,China;2.College of Life Sciences,Shanxi University,Taiyuan 030006,China)
The project was planned to start from sclerotia differentiation,effect of exogenous signal molecule on sclerotial differentiation of Penicillium thomii PT95 was studied.Using MEA and PDA two different culture medium of Penicillium thomii PT95, and 0,0.02,0.04,0.06 mmol/mL of cAMP separately on two different media,by recording the time to sclerotia mature,measuring the colony diameter,sclerotia biomass and carotenoid content,to analyze the growth and development of the strains.The results showed that when the cAMP concentration reached 0.02 mmol/mL,the growth state of sclerotium was the best;after increases cAMP concentrations growth status decreased;when the cAMP concentration reached 0.06 mmol/mL,the growth began to be suppressed.That indicated that under the low concentration of cAMP,Penicillium thomii PT95 strain's growth was promoted.But if the supply of cAMP concentration was toohigh,sclerotia biomass and carotenoid content ofPT95 strain would be lower than normal,the growth was inhibited.
cAMP;Penicillium thomii;sclerotia;carotenoids
Q935
A
1002-2481(2016)01-0019-05
10.3969/j.issn.1002-2481.2016.01.06
2015-07-30
国家自然科学基金项目(31070048)
赵文婧(1980-),女,山西榆次人,讲师,博士,主要从事微生物生理生化研究工作。韩建荣为通信作者。
