马铃薯幼苗低温胁迫的生理响应及品种耐寒性综合评价
2016-12-17杨慧菊郭华春
杨慧菊,郭华春
(云南农业大学农学与生物技术学院薯类作物研究所,云南 昆明 650201)
马铃薯幼苗低温胁迫的生理响应及品种耐寒性综合评价
杨慧菊,郭华春*
(云南农业大学农学与生物技术学院薯类作物研究所,云南 昆明 650201)
以马铃薯品种‘宣薯2号’为试验材料,采用人工模拟低温的方法,对马铃薯幼苗离体叶片进行不同条件的低温胁迫。低温处理后的叶片,分别测定其叶绿素含量、相对电导率、POD活性、SOD活性、MDA含量、游离脯氨酸含量、可溶性糖含量和可溶性蛋白含量的变化,通过相关性分析,筛选出评价马铃薯苗期耐寒性的关键生理生化指标。结果表明,随着温度的降低和低温胁迫时间的延长,马铃薯幼苗相对电导率逐渐增大;叶绿素含量逐渐降低;POD活性和SOD活性总体上表现为先急剧升高后降低,再缓慢升高的变化;MDA含量呈先急剧增加,后下降的变化;游离脯氨酸和可溶性糖含量含量均表现为先升后降的变化;可溶性蛋白含量呈现不断上升的变化。相对电导率、叶绿素含量、POD酶活性、SOD酶活性、可溶性糖含量、可溶性蛋白含量各指标间都达到了显著或极显著相关水平。为了更全面准确的评价马铃薯品种耐寒性,用隶属函数法综合这些指标,对14个马铃薯品种(系)苗期耐寒性进行综合评价,其耐寒性从强到弱排序依次为转心乌>会-2>冀张薯12号>丽薯6号>中薯18号>晋薯16号>米拉>费乌瑞它>滇同薯1号>滇薯716>芒果洋芋>宣薯2号>合作88>威芋3号。
马铃薯;幼苗;低温胁迫;生理响应;隶属函数
马铃薯(SolanumtuberosumL.)是世界上仅次于水稻、玉米和小麦的第四大粮食作物。经过近几十年来的研究与开发,以马铃薯为原料的加工产品得到了快速发展,目前全世界马铃薯加工产品不断增加[1-3]。在中国南方利用冬季闲田种植马铃薯,是近年马铃薯种植面积不断增加的重要因素,且冬闲稻田生产马铃薯在销售价格、资源利用、栽培方式和环境保护等方面具有较多优势[4]。发展冬马铃薯,低温是影响马铃薯生长及产量的重要环境因子,特别是0 ℃以下的低温冻害会对马铃薯植株的叶片、茎秆造成严重的损伤,甚至导致整个植株的死亡[5-6]。马铃薯栽培种虽喜冷凉,但不耐霜冻,有研究表明,温度低于1 ℃时马铃薯植株停止生长、-0.8 ℃时遭受冷害、-1.5 ℃时遭受冻害、-3 ℃时植株会冻死[7]。在中国,冬季低温会对马铃薯造成霜冻危害,早春种植的马铃薯在苗期也常遭倒春寒造成减产,秋季种植的马铃薯在块茎膨大期遭受低温、早霜或晚霜的影响,使块茎生长受阻[8],导致减产。
Chen等利用Steffen的方法,在0~-7 ℃内测定不同种的马铃薯叶片在各个温度时的电导率,最后算出各个材料的致死温度,用于评价各个材料的耐冻性[9]。质膜透性越大,电导率就愈大,致死温度越高,耐冻性越差[10-13]。辛翠花[10]等研究表明,低温胁迫下SOD 活性呈现出先降后升再降的变化规律,而POD的活性表现出先升后降。除上述几个抗寒性生理生化指标外,叶绿素含量也可作为抗寒性鉴定指标,温度过高和过低都会抑制酶反应,甚至会破坏原有的叶绿素[14]。Levitt等研究表明,低温驯化期间,马铃薯糖含量增加的同时伴随着抗寒性的增强[15]。Chen和Li研究发现,通过低温驯化后的马铃薯叶片含有较高水平的可溶性蛋白,且可溶蛋白的增加与抗寒性成正相关[16]。李飞等以低温敏感的中薯3号和抗寒性较强的野生马铃薯03079-435(Solanumacaille)幼苗为试材,研究了驯化期间的MDA含量变化,发现中薯3号的MDA整体含量都高于野生种,表明了抗寒性差的马铃薯在低温胁迫下膜损伤程度较大,受霜冻影响更为严重[17]。
本研究通过人工模拟低温环境的方法,在马铃薯生长的苗期对其进行低温胁迫,通过测定其叶片各生理生化指标含量的变化,及其相关性分析,筛选出评价马铃薯抗寒能力的关键性指标,为马铃薯苗期抗寒性评价提供理论依据。运用隶属函数法综合评价不同马铃薯材料的抗寒性,为冬季和早春季节马铃薯种植品种的选择奠定理论基础,提高马铃薯种植效益,并能为马铃薯抗寒育种中亲本材料的选择提供依据。
1 材料与方法
1.1 试验材料与培养
试验于2013年在云南农业大学薯类作物研究所进行,供试的14个马铃薯品种(系):会-2、宣薯2号、合作88、费乌瑞它、丽薯6号、芒果洋芋、滇同薯1号、晋薯16号、威芋3号、冀张薯12号、滇薯716、中薯18号、转心乌、米拉。分别取大小均匀的块茎,用10 mg/L的赤霉素催芽,20 min取出晾干,待发芽后,取芽长一致的种薯,种植于10 cm×12 cm的营养钵中,1钵1粒(培养基质为营养土与蛭石按1∶1比例混合),置于植物气候箱中培养(温度18~20 ℃,光照强度50~60 μmol/m2·s,相对湿度65 %±10 %)。
1.2 试验设计与处理
待幼苗生长至20 cm(苗龄4周)时,分别取倒4叶片,用蒸馏水冲洗干净,吸水纸将叶片表面水分擦干,放入人工低温恒温槽中,进行不同条件的低温胁迫。低温胁迫的温度梯度为-2、-4、-6 ℃,低温胁迫的时间梯度为1,3,6,9,12 h。低温胁迫处理后的叶片,混合均匀,进行各项生理生化指标的测定,每个指标重复4次。同时以正常生长条件(20 ℃)为温度胁迫对照,0 h为时间胁迫对照。
1.3 测定指标与方法
相对电导率(REC)采用外渗电导法;叶绿素(Chl)含量采用分光光度法;过氧化物酶(POD)活性采用愈创木酚法;超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法;丙二醛(MDA)含量采用硫代巴比妥酸比色法;游离脯氨酸(Pro)含量采用磺基水杨酸法;可溶性糖(SS)含量采用蒽酮比色法;可溶性蛋白(SP)含量采用考马斯亮蓝G-250法,以上方法均参照张宪政主编的《作物生理研究方法》[18]进行测定。
1.4 品种耐寒性综合评价方法
采用模糊综合评判方法中的隶属度公式,对试验原始数据进行定量转换,以综合评价不同马铃薯品种(系)的耐寒性。隶属函数值的计算参考杨金红[19]的方法,具体步骤如下:如果某一指标与耐寒性呈正相关,则公式为:X(u)=(X-Xmin)/(Xmax-Xmin),如果某一指标与耐寒性呈负相关,则公式为:X(u)=1-(X-Xmin)/(Xmax-Xmin)。
其中,X(u)为各指标的隶属度值;X为某一指标的测定值;Xmin为某一指标测定值中的最小值;Xmax为某一指标测定值中的最大值。计算各指标隶属函数值的平均值用隶属函数法评定时,隶属函数平均值越大,则耐寒性越强。
1.5 数据统计与分析
采用Microsoft Excel和SPSS 18.0 软件进行数据处理、差异显著性分析和作图。
2 结果与分析
2.1 不同低温胁迫下宣薯2号叶片中各生理生化指标的变化
2.1.1 不同低温处理下叶绿素含量的变化 如图1所示,不同温度胁迫下,随着温度的降低,马铃薯幼苗叶片中叶绿素总含量(Chl)、叶绿素a(Chla)含量和叶绿素b(Chlb)含量均下降。-2 ℃胁迫下,各叶绿素含量下降明显,-2 ℃时,下降较缓慢。Chla/Chlb随着温度的降低,先急剧升高,-4 ℃时,缓慢升高。说明Chl、Chla和Chlb含量的变化趋势一致,即低温对3种叶绿素含量的分解具有相同程度的影响,分解速率变化趋势一致。
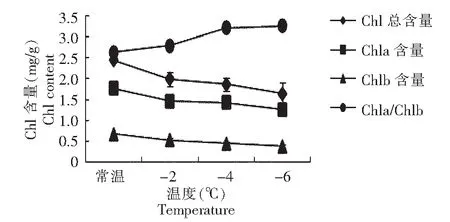
图1 不同温度胁迫下叶绿素含量的变化Fig.1 Changes of chlorophyll content under different low temperature stress
由图2可看出,不同时间-2 ℃低温胁迫条件下,随着时间的延长,Chl、Chla和叶Chlb含量均大幅度下降,1~6 h各叶绿素含量均升高,6~9 h下降,9~12 h又升高,呈现先升后降,再升高再降低的变化。Chla/Chlb随着低温胁迫时间的延长,先升高,3 h时达到最大值,3 h后开始下降,6 h以后再升高,9~12 h升高较为迅速。表明低温胁迫对Chla和Chlb破坏程度不同,Chla/Chlb升高,说明Chla降解速率小于Chlb,低温对Chla的破坏程度小于Chlb;Chla/Chlb下降,说明Chla降解速率大于Chlb,低温对Chla的破坏程度大于Chlb。
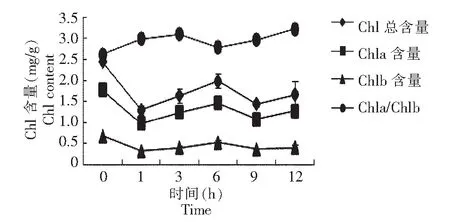
图2 不同时间低温胁迫下叶绿素含量的变化Fig.2 Changes of chlorophyll content under low temperature different time stress
2.1.2 不同温度处理下各生理生化指标的变化 由表1可看出,不同低温胁迫下,随着温度的逐渐降低,马铃薯幼苗叶片中相对电导率呈增大趋势,其中-4 ℃胁迫后增大较为迅速;POD活性不断上升,变化幅度较大,在-2~-4 ℃,POD活性急剧上升,之后上升缓慢。SOD活性先缓慢上升,到-4 ℃下降,之后又急剧上升,-6 ℃达最大值;随着温度的降低,MDA含量呈先急剧增加,后下降的变化,其中在-2 ℃时达到最大值;游离脯氨酸含量先急剧上升,-4 ℃时达最大值,-4 ℃后又下降,总体表现为先升后降的变化趋势;可溶性糖含量先迅速升高,-2 ℃时达到最大值,-2 ℃后缓慢下降,总体表现为先升高后降低的变化趋势;可溶性蛋白含量先缓慢升高,-4 ℃急剧升高,-6 ℃达最大值,总体表现为不断升高的变化趋势。
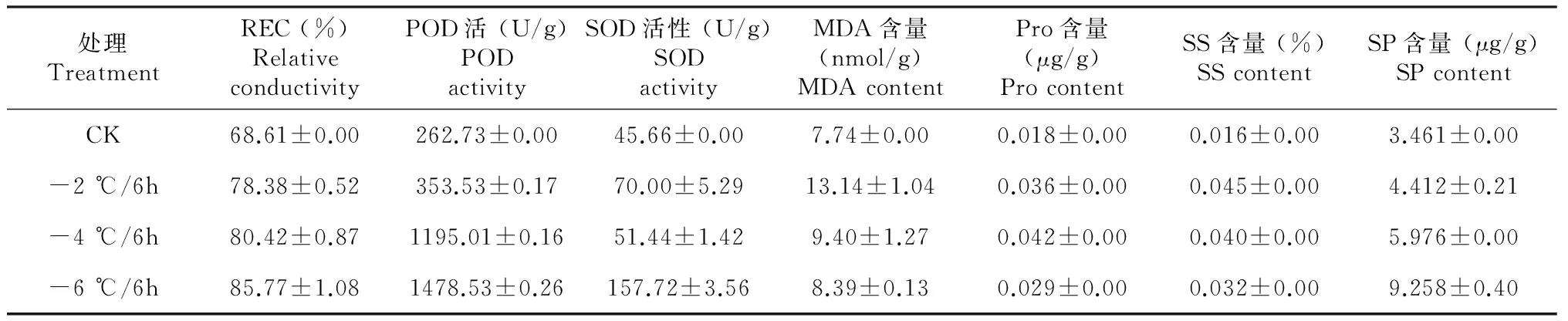
表1 不同温度胁迫下各生理生化指标的变化
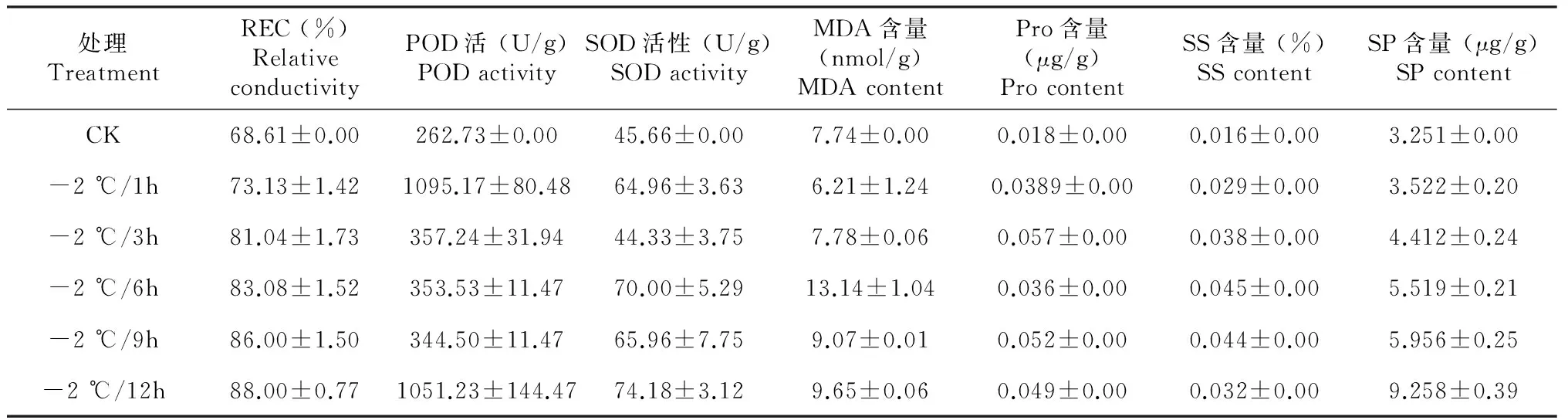
表2 不同时间低温胁迫下各生理生化指标的变化
2.1.3 低温处理不同时间各生理生化指标的变化 由表2可知,-2 ℃低温胁迫下,随着胁迫时间的延长,马铃薯幼苗叶片中相对电导率不断增加,其中胁迫0~3 h增幅较大;POD活性急剧上升,胁迫1 h达到最大值,之后迅速下降,其中在6~9 h呈缓慢下降,接近低温胁迫前的水平,9 h开始上升,POD活性总体上表现出急剧上升后下降,再上升的变化趋势;SOD活性表现为先急剧升高后降低,再缓慢升高的趋势,其中胁迫6 h时达最大值;MDA含量先缓慢下降,胁迫1 h大幅度上升,6 h达到最大值,6 h后又下降;游离脯氨酸含量先升高,胁迫3 h达最大值,3 h后又下降,胁迫6 h以后变化趋势与0~6 h变化一致,均为先升高后降低;随着胁迫时间的延长,可溶性糖含量先缓慢升高,胁迫6 h达最大值,6 h后呈下降趋势,胁迫12 h时可溶性糖含量仍高于胁迫前;可溶性蛋白的含量先缓慢升高,胁迫9 h时迅速上升,胁迫12 h时达最大值,表现为不断升高的变化趋势。
2.1.4 马铃薯苗期耐寒性指标之间的相关性分析 根据表3中各指标间的相关性分析可知,过氧化物酶活性与相对电导率呈极显著负相关,相关系数为r=-0.600**;过氧化物酶活性与叶绿素含量呈显著正相关,相关系数为r= 0.558*;超氧化物歧化酶活性与相对电导率呈显著负相关,相关系数为r=-0.442*;超氧化物歧化酶活性与叶绿素含量呈显著正相关,相关系数为r= 0.524*;可溶性糖含量与叶绿素含量呈显著正相关,相关系数为r= 0.491*;可溶性糖含量与过氧化物酶活性呈显著正相关,相关系数为r= 0.524*;可溶性蛋白含量与超氧化物歧化酶活性呈显著正相关,相关系数为r= 0.558*。丙二醛含量和游离脯氨酸含量与其它指标间的相关性均未达显著或极显著水平。
2.2 低温胁迫下不同马铃薯品种(系)叶片中生理生化指标的变化
2.2.1 不同马铃薯品种(系)低温胁迫后生理生化指标的变化 由表4可知,转心乌叶片中相对电导率增幅最小,为60.63 %,米拉相对电导率增幅最大,为78.96 %,说明米拉叶片细胞膜损伤程度较重,电解质渗出率变化较大;转心乌叶片中叶绿素含量降幅最小,为0.0706 mg/g,威芋3号叶片中叶绿素含量降幅最大,为0.6725 mg/g,说明其叶片细胞造成了严重损伤,使叶绿素大量降解;转心乌叶片中POD活性和SOD活性增幅均最大,分别为1060.89和44.82 U/g,而威芋3号叶片中POD活性和SOD活性增幅均最小,分别为340.69和7.81 U/g,说明转心乌保护酶系统清除活性氧和自由基伤害的能力较强,而威芋3号较弱;威芋3号叶片中MDA含量的增幅最大,为2.34 μmol/g,转心乌叶片中MDA含量增幅最小,为0.35 μmol/g,说明威芋3号膜质过氧化程度大,叶片中MDA积累量多;转心乌叶片中游离脯氨酸的增幅最大,为0.3087 μg/g,威芋叶片中3号游离脯氨酸的增幅最小,为0.0348 μg/g;转心乌叶片中可溶性糖和可溶性蛋白增幅均最大,分别为0.9438 μg/g和0.17 %,合作88叶片中可溶性糖含量增幅最小,为0.01 %,威芋3号叶片中可溶性蛋白含量增幅最小,为0.5742 μg/g。
2.2.2 不同马铃薯品种(系)耐寒性综合评价 由表5可得,14个马铃薯品种(系)各指标的隶属函数平均值(耐寒性大小)从大到小顺序依次为转心乌>会-2>冀张薯12号>丽薯6号>中薯18号>晋薯16号>米拉>费乌瑞它>滇同薯1号>滇薯716>芒果洋芋>宣薯2号>合作88>威芋3号。用隶属函数法对植物进行抗逆性评价时,隶属函数平均值越大,其耐寒性越强,由此可得,14个马铃薯品种(系)中,耐寒性最强的是转心乌,最弱的是威芋3号。
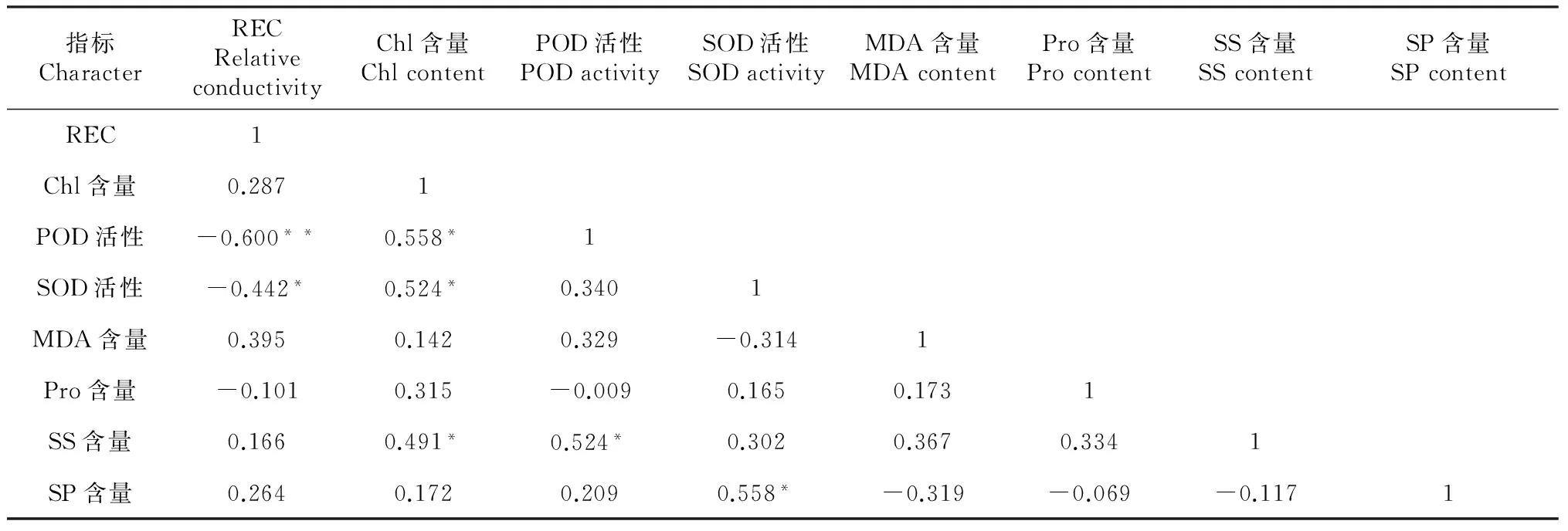
表3 马铃薯幼苗耐寒性指标间的相关性
注:r*为0.05水平上显著,r**为0.01水平上显著。
Notes:r*andr** meant significantce at 5 % and 1 % levels, respectively.

表4 低温胁迫下14个马铃薯品种(系)生理生化指标的变化
注:小写字母表示0.05水平差异显著,下同。
Notes:Lowercase shows significant difference at 0.05 level. The same as below.
3 讨 论
在评价植物耐寒性的生理生化指标中,质膜电解质渗出率即相对电导率的大小是植物抗寒力强弱的重要指标[20],可用来判断植物受低温伤害程度和植物的抗寒能力。本研究中,随着温度的降低和低温胁迫时间的延长,相对电导率均不断增加,与魏亮等[21]的研究结果相符合。叶绿素是植物进行光合作用的主要条件之一,在一定程度上可以反应植物的生产性能和抵耐逆境胁迫的能力[22]。本研究中随着温度的降低和低温胁迫时间的延长,马铃薯叶片中总叶绿素含量逐渐降低,Chla/Chlb随着温度的降低逐渐升高,但随着低温胁迫时间的延长,Chla/Chlb先升高,后下降。有研究表明,植物叶绿素含量的变化是通过Chla和Chlb减少产生的,Chla/Chlb比值增大,表明在轻度的低温胁迫下,降低了对光能捕获和传递,但加强了光合反应中心对光能的转换,当超过一定温度和时间范围时,就会引起植株体内大量积累自由基和活性氧,特异性地破坏Chla,使Chla/Chlb下降[23-24]。
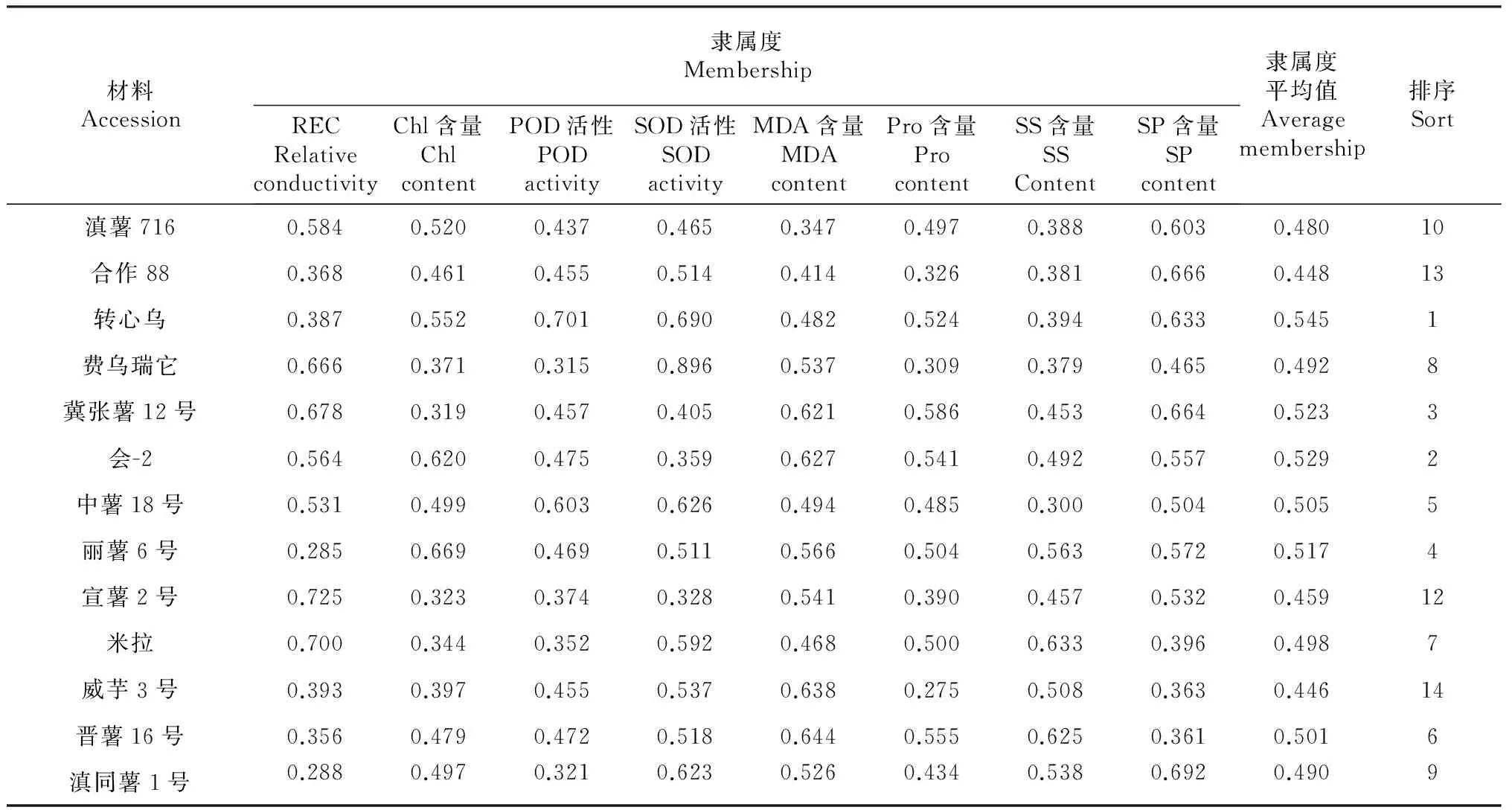
表5 各指标隶属函数值及耐寒性综合评价
植物经低温处理后体内的多种酶活性产生明显变化,酶活性提高和降低与植物抗寒性的强弱呈一定相关性[25]。本研究中随着温度的降低,马铃薯幼苗叶片中SOD、POD活性不断增加,且增幅较大,与前人研究结果一致。
MDA是膜质过氧化的终产物之一,其含量多少代表质膜受损伤的程度,因此也可作为膜系统受伤害的指标之一[26]。由于MDA含量的测定简单快速,目前在抗寒性研究中应用较广,但仍有很多研究表明,MDA含量不能作为判断植物受损伤严重与否的指标,因为MDA含量的测定容易受其它因素的干扰[27-28]。本研究中,低温胁迫下马铃薯叶片中MDA含量差异达到显著水平,但含量较低,且MDA含量与其它指标间的相关性均未达显著水平,这可能与MDA含量的测定方法,或离体叶片材料的选取有关。
游离脯氨酸的积累与植物抗寒性的关系表现为两种情况:在某些情况下,游离脯氨酸含量的增加可以提高植物抗寒力;但在另一些情况下,则不显示出有任何的相关性[29]。李飞等[17]研究表明,霜冻敏感的中薯3号在整个冷驯化期间脯氨酸含量一直呈下降趋势,在后期又略有升高,整个驯化期间无显著差异。Van等研究发现,马铃薯植株脯氨酸含量与抗寒性之间具有高度相关性,在低温胁迫下,抗寒性越强的植株,脯氨酸含量越高[30]。本研究结果表明,游离脯氨酸积累量与马铃薯耐寒性有一定关系,但与其它各耐寒性指标间相关性不显著。
Levitt等[15]研究发现,低温驯化期间,马铃薯糖含量增加的同时伴随着抗寒性的增强,本研究结果与之相符,说明马铃薯叶片中可溶性糖的积累量与其耐寒性有密切关系。可溶性蛋白含量的增加,与抗寒性的增强相一致,抗寒锻炼中的蛋白质代谢,对抗寒性的提高担负着重要作用,可溶性蛋白不但在量上增加,在质上也有变化[31]。Chen和Li研究发现,通过低温驯化后的马铃薯叶片含有较高水平的可溶性蛋白,野生种马铃薯的叶片中可溶蛋白的增加与抗寒性成正相关[16],本研究表明,在低温胁迫下,马铃薯叶片中可溶性蛋白含量逐渐增加,说明可溶性蛋白含量与马铃薯耐寒性成正相关。
本研究通过各指标综合分析表明,MDA含量和游离脯氨酸含量与其它指标都无显著性或极显著的相关性,因此,在利用MDA含量和游离脯氨酸含量来鉴定马铃薯幼苗的耐寒性必须结合其它指标综进行综合评价才合理。隶属函数法已经成功应用于葡萄[32]、玉米[33]等作物的抗逆性评价,本研究应用隶属函数分析法马铃薯苗期的耐寒性,以8个生理生化指标对不同马铃薯品种进行综合耐寒性评价,提高了耐寒性评价的准确性。
采用耐寒性室内评价的方法,能够人为控制植物的生长条件,使植株低温胁迫的温度和时间一致,能够更准确、可靠地对植物进行耐寒性鉴定。但本研究采用的是马铃薯离体叶片,通过叶片所测得的耐寒性与其整个植株的耐寒性之间可能存在差异。
[1]屈冬玉, 谢开云, 金黎平, 等. 中国马铃薯产业发展与食物安全[J].中国农业科学, 2005, 38(2):358-362.
[2]李燕山, 姚春光, 普红梅,等. 不同施肥模式对马铃薯云薯401生长及产量的影响[J]. 南方农业学报, 2014, 45(9):1608-1611.
[3]于健龙, 杨永奎, 胡 辉,等. 不同栽培措施及肥料配比对马铃薯农艺性状及产量的影响[J]. 南方农业学报, 2014, 45(7):1215-1220.
[4]谢开云, 王晓雪, 王凤义. 我国冬闲稻田马铃薯生产的潜力与问题[J].中国马铃薯,2010, 24(6):370-375.
[5]魏 亮, 李 飞, 徐建飞, 等. 马铃薯抗寒性研究进展[J].贵州农业科学, 2012, 40(2):44-47.
[6]李国璋, 温芸芸. 宁夏隆德县马铃薯全生育期气象条件分析[J]. 安徽农业科学, 2015(34):294-295.
[7]门福义, 刘梦芸. 马铃薯栽培生理[M]. 北京:中国农业出版社, 1995.
[8]李 芳. 马铃薯耐霜冻能力鉴定以及低温处理下表达谱分析[D]. 华中农业大学博士学位论文, 2011.
[9]吴能表, 钟永达, 肖文娟, 等. 零上低温对甘蓝幼苗逆境指标的动态影响[J]. 西南师范大学学报, 2005, 30(3):525-528.
[10]徐 康, 夏宜平, 徐碧玉, 等. 以电导法配合Logistic方程确定茶梅“小玫瑰”的抗寒性[J]. 园艺学报, 2005,32(1):148-150.
[11]Vega S E, Rio A H, Jung G. Marker-assisted Genetic Analysis of Non-acclimated Freezing Tolerance and cold Acclimation Capacity in a BackcrossSolanumPopulation[J]. Amer J of potato Res, 2003, 80:59-369.
[12]Nunes M E S, Smith G R. Electrolyte Leakage Assay Capable of Quantifying Freezing Resistance in Rose Clover[J].Crop Science, 2003, 43:1349-1357.
[13]辛翠花, 蔡 禄, 肖欢欢, 等. 低温胁迫对马铃薯幼苗相关生化指标的影响[J]. 广东农业科学, 2012(22):19-21.
[14]Nagata N, Tanaka R, Satoh S. Identification of a vinyl reductase gene for chlorophyⅡ synthesis in Arabidopsis thaliana and implication for the evolution of Prochlorococcus specie[J]. Plant Cell, 2005, 17(1):233-240.
[15]Levitt J. Responses of Plants to Environmental Stresses:Volume 1, Chilling, freezing and high temperature stresses[J]. Physiological ecology, 1980.
[16]Chen T H H, Gusta L V. Abscisic acid-induced freezing resistance in cultured plant cells[J]. Plant Physiology, 1983, 73(1):71-75.
[17]李 飞, 刘 杰, 段绍光, 等. 马铃薯幼苗在冷驯化期间的生理生化变化[J].中国马铃薯, 2008, 22(5):257-260.
[18]张宪政.作物生理研究方法[M]. 北京:农业出版社,1992.
[19]杨金红. 山东地区引种石楠属植物苗期生长和抗寒性研究[D]. 南京林业大学, 2007.
[20]张昕欣. 腊梅品种抗寒性研究[D]. 南京林业大学, 2008.
[21]魏 亮. 中国主要马铃薯品种抗寒性鉴定及抗寒相关基因表达分析[D]. 内蒙古农业大学, 2012.
[22]张明生, 谈 锋, 张启堂. 快速鉴定甘薯品种耐旱性的生理指标及方法的筛选[J]. 中国农业科学,2001,34(3):260-265.
[23]李小燕, 丁丽萍. 自然越冬状态下四翅滨藜抗寒性生理指标的动态变化[J]. 东北林业大学学报, 2008,36(5):11-12.
[24]伍泽堂. 超氧自由基与叶片衰老时叶绿素破坏的关系[J]. 植物生理学通讯,1991,27(4):277-280.
[25]彭永康, 郝泗城, 王振英. 低温处理对豇豆幼苗生长和 POD、COD、ATPase同功酶的影响[J]. 华北农学报, 1994,9(2):76-80.
[26]艾希珍, 于贤昌, 王绍辉, 等. 低温胁迫下黄瓜嫁接苗与自根苗某些物质含量的变化[J]. 植物生理学通讯, 1999,35(1):26-29.
[27]DraPer H H, Hadley M. Malondialdehyde determination as index of lipid peroxidation[J]. Methods Enzymol, 1990,21(1):33-37.
[28]Vick B A, Zimmerman D C. Metabolism of fatty acid hydroperoxides by Chtorella pyrenoidosa[J]. Plant Physiology, 1989,90(1):125-132.
[29]杨凤翔. 不同草莓品种抗寒性的综合评价[D]. 甘肃农业大学, 2010.
[30]Van Swaaij A C, Jacobsen E, Feenstra W J. Effect of cold hardening, wilting and exogenously applied praline on leaf praline content and frost tolerance of several genotypes ofSolanum[J]. Physiology Plantarum, 1985, 64:230-236.
[31]张石城. 植物的抗寒生理[M]. 北京:农业出版社, 1990.
[32]张文娥, 王 飞, 潘学军. 应用隶属函数法综合评价葡萄种间抗寒性[J]. 果树学报, 2007,24(6):849-853.
[33]许桂芳, 张朝阳, 向佐湘. 利用隶属函数法对4种珍珠菜属植物的抗寒性综合评价[J]. 西北林学院学报, 2009, 24(3):24-26.
(责任编辑 王家银)
Physiological Responds of Potato Seedlings to Low Temperature Stress and Comprehensive Evaluation on Their Cold Tolerance
YANG Hui-ju,GUO Hua-chun*
(Tuber-Root Crops Research Institute, College of Agronomy and Biotechnology, Yunnan Agriculture University, Yunnan Kunming 650201,China)
Using potato variety ‘Xuanshu 2’ as materials, we studied the changes of relative conductivity, content of chlorophyll, POD activity, SOD activity, contents of MDA, free proline, water soluble sugar and water soluble protein under different low temperature stress by artificial, at the seedling stage of potatoes. Based on these physiological and biochemical indexes correlation analysis, Some key indexes were screened out which could evaluate the cold tolerance of potato seedlings, the results indicated that with the lowering of stress temperature and the prolonging of stress time, the relative conductivity of potato seedlings was increased, content of chlorophyll was decreased, POD activity and SOD activity were firstly increased and then decreased, increased slowly at last, content of MDA, free proline and water soluble sugar were firstly increased and then decreased, content of water soluble protein was constantly increased. The relative conductivity, content of chlorophyll, POD activity, SOD activity, water soluble sugar and water soluble protein, were significant or extremely significant respectively, but a single index could not evaluate the cold tolerance of potato seedlings. It must be combined with other indexes to a comprehensive evaluation. Evaluation on cold tolerance of 14 potato varieties were made through fuzzy comprehensive evaluation method, the cold tolerance order was Zhuanxinwu>Hui-2>Jizhangshu12>Lishu6>Zhongshu18>Jinshu16>Mira>Favorita>Diantongshu1>Dianshu716>Mangguoyangyu>Xuanshu2>Hezuo88>Weiyu3.
Potato; Seedling; Low temperature stress; Physiological respond; Membership function
1001-4829(2016)11-2560-07
10.16213/j.cnki.scjas.2016.11.009
2015-01-06
国家马铃薯产业体系 (GARS-10);国家自然科学基金(C130407);云南省重大科技专项(2013ZA007)
杨慧菊(1989-),博士研究生,主要从事马铃薯生理生态学研究,E-mail:851850213@qq.com,*为通讯作者,E-mail:ynghc@126.com。
S532
A
