早古生代烃源形成的生物组合及其意义
2016-11-22刘文汇腾格尔卢龙飞谢小敏
刘文汇,胡 广,腾格尔,王 杰,卢龙飞,谢小敏
(1.中国石化 石油勘探开发研究院 无锡石油地质研究所,江苏 无锡 214151;2.西南石油大学 地球科学与技术学院,四川 成都 610500)
早古生代烃源形成的生物组合及其意义
刘文汇1,胡 广2,腾格尔1,王 杰1,卢龙飞1,谢小敏1
(1.中国石化 石油勘探开发研究院 无锡石油地质研究所,江苏 无锡 214151;2.西南石油大学 地球科学与技术学院,四川 成都 610500)
早古生代生物的富集和埋藏为油气资源提供了雄厚的物质基础。不同生物对烃类贡献的差异使成烃生物组合成为烃源岩研究的重要内容,也是高演化烃源岩评价的一个新手段。从地质历史生物演化来看,藻类是早古生代最主要的成烃生物先质,按照生活习性,它们可以分为底栖藻类和浮游藻类。底栖藻类主要生活在海岸带,成带状分布,主要受光的强度、基底情况等因素控制;浮游藻类主要生活在表层水体,它们在不同海域的分布主要受藻体大小和风浪强度控制。依据对塔里木盆地北缘4条下古生界露头剖面的7个地层组中成烃生物的辨识及有机质地球化学特征的研究,总体显示以底栖藻类为主的烃源岩的有机碳含量高于以浮游藻类为主的烃源岩,除形成环境的因素外,成源和生烃过程均有影响,但后者对有机质的碳同位素值影响较小。烃源岩干酪根碳同位素值(δ13C干酪根)与成烃生物组合面貌对比研究发现,以底栖藻类为主的烃源岩的δ13C干酪根小于-34‰;以浮游藻类为主的烃源岩的δ13C干酪根大于-30‰。对比塔里木盆地已发现原油的碳同位素值(均重于-30‰),推断其原油主要来源与浮游藻类密切相关,揭示了下古生界具有较低有机碳含量但以浮游藻类为主的源岩具有很大的生烃能力。
成烃生物;有机碳含量;干酪根碳同位素值;油-源对比;早古生代;塔里木盆地
古生界海相碳酸盐岩层系成为中国油气勘探的重要领域。塔里木盆地下古生界大型油田和四川盆地古生界大型气田的发现,彰显了古生界油气的巨大潜力。遗憾的是,按照目前对烃源的认识和油气源的对比,还无法确定这些油气的直接源岩。传统认识的优质烃源,其分布和规模难以提供目前已探明的油气资源。如何认识古生界的烃源岩,直接影响油气勘探的部署和对油气潜力的认识。经典烃源评价的有机地球化学指标,无法应对时代老、演化程度高、叠加的地质作用造成的地球化学复杂、过程恢复困难的古老烃源。为此,部分学者从生物的角度对高演化烃源岩生烃潜力评价已开展了一些探索性的工作[1-4]。他们的研究成果为古生代烃源研究工作打下了基础,开拓了思路。
近年来对现代生物生烃热模拟和成烃生物的研究相继积累了一些基础资料,形成了一些创新成果,诸如吴庆余等(1987)对蓝藻的热模拟实验讨论了浮游藻类的产气能力及产物特征[5];张惠之等(1994)针对陆相地层中出现的盘星藻化石进行了生烃模拟试验[6];宋一涛等(1995)研究了浮游藻类的生烃能力及产物特征,包括生物标志物化合物的特征;李超等[7](2001)模拟了单细胞海藻的生烃过程[8];郭汝泰和杨凤丽(2002)研究了65℃条件下多细胞马尾藻对低熟油的生烃贡献[9];刘文斌等(2005)研究了海带的热演化程度对排油过程的影响[10];叶云和李玉成等(2012)讨论了巢湖蓝藻腐殖化过程中的形态与成分变化[11]。这些研究已初步揭示了不同生物类型有机质生烃潜力的明显差异。
由此可见,从生物的角度对烃源进行研究可能是对高演化烃源岩评价的一个突破口。然而,由于下古生界形成年龄老、演化时间长、生物原始面貌辨识困难等原因,目前关于早古生代成烃生物的生物构成、生态分布及含不同成烃生物组合烃源岩的地球化学特征还缺乏系统研究。我们从地质历史生物演化序列入手,刻画早古生代主要成烃生物的构成及生态分布,并以塔里木盆地北缘古生界沉积有机质为重点对象,探讨早古生代不同成烃生物烃源岩的地球化学特征,进而对塔里木盆地下古生界海相碳酸盐岩层系进行油-源对比研究。
1 早古生代成烃生物
成烃生物是指生物群中可为生烃作贡献的那部分生物[12],并不等同于地层中的古生物群。与此类似的概念,例如“生烃母质”在1984年Tissot就已提出[13]。因此,成烃生物是把烃源岩中的生物碎片从生油生物整体来研究的,与古生物学中的生物群和有机岩石学中的显微组分均有一定的差别。依据现代生烃模拟实验,最重要的成烃生物主要是由藻类组成[3,5-11]。
最早的光自养藻类至少在2 700 Ma就存在于海洋中[14],大量有机质壁真核浮游藻类存在于1 600~1 800 Ma的地层中[15]。Butterfield(2000)对保存形态较好的有机质壁化石研究认为,红藻起源于约1 200 Ma[16]。到新元古代,部分浮游绿藻已被证实为真核藻类。而中生代和现代海洋中,藻类占主要地位的是真核藻类。
早古生代有机质壁浮游藻类显著多样化分化与寒武纪—奥陶纪海洋无脊椎生物大辐射平行。其演化按色素系列可分为“绿色色素演化序列”和“红色色素演化序列”。其中,“绿色色素演化系列”藻在190Ma之前占海洋藻类的主体。留下比较清晰形态学证据的古生代浮游绿藻是塔斯马利亚藻,该藻类化石在整个古生代都存在,在二叠纪达到高峰,并在塔斯马尼亚地区和阿拉斯加地区沉积了所谓的“白煤”和油页岩。具有“红色色素演化系列”特色的浮游藻类在新元古代—古生代沉积序列中就有记录,例如Moldowan(1998)爱沙尼亚塔林地区下寒武统Lukati组含大量疑源类页岩中检出甲藻所特有的生物标志化合物重排甾烷和4-甲基-24-乙基胆甾烷[17]。
早古生代的成烃生物主要包括细菌、浮游藻类、底栖宏观藻类和动物有机质[1]。晚奥陶世—早志留世植物登陆之后,成烃生物还包括类似苔藓类植物。此外在晚元古代—志留纪,还有一类植物界于藻类和高等植物之间,被称之为线叶植物[1]。按生物的演化,早古生代对生烃有贡献的生物主要包括蓝藻、单细胞藻、多细胞藻、真细菌、古生菌、真菌、甲藻和苔藓植物(图1)[18]。
藻类作为早古生代最主要的成烃生物,按其生活习性可以分为底栖藻类和浮游藻类。如前所述,底栖和浮游藻类的生烃能力和特征明显不一致,因此区分底栖藻类和浮游藻类对烃源岩的评价意义重大。
对现代藻类生活习性的研究发现,底栖藻类主要有褐藻和红藻,还有部分为绿藻门的石莼纲。虽然部分红藻和褐藻的生活水深最大可以达到200 m左右,但绝大多数底栖藻类主要还是生活在海岸带。这些海岸生的藻类,构成鲜明的植物群落,它们成带分布的最主要决定因子是海浪、基质的情况和光的强度。一般在40~60 m的水深带内是以光谱中的短波部分的蓝绿光线占主要优势,对多数红藻是非常适宜的,而对其他藻类,要达到最大程度的同化作用的环境,则是很不适应的。
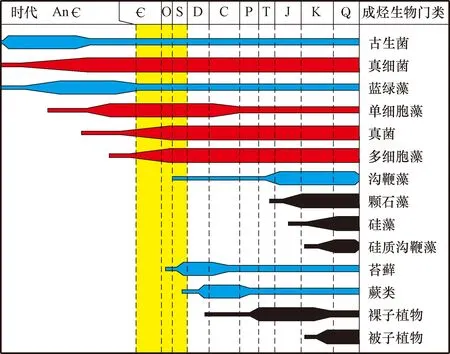
图1 沉积有机质的主要贡献[18]Fig.1 Contributions from deposited organic matters
现代海洋的浮游植物主要是蓝藻、硅藻和双鞭毛藻(甲藻)。相对来讲,硅鞭毛藻和绿鞭毛藻较少,裸藻和黄藻更少。对于古生代海洋而言,已有的不完整的研究表明占主导地位的浮游藻类主要有蓝藻、粘球型藻、塔斯马尼亚藻和疑源类。与底栖藻类类似,在不同的海域,浮游藻类的组合量及其属种也不一样,但主要控制因素为藻体大小、营养组分和风浪强度。浮游藻类按其个体大小可分为超微型浮游藻类和大型浮游藻类。超微型浮游藻类主要是由微球状蓝藻组成(大小稍大于2 μm,到5 μm左右),主体是球状蓝藻聚球藻(synechococcus)、聚胞藻(synechocystis)和原绿球藻。由于这些超微型浮游藻类个体小,具有高的比表面积,加上在微小细胞周围高的扩散梯度,允许细胞高速地摄取营养。大型浮游藻类个体较大,藻类细胞中产生伪空泡,依靠这些伪空泡聚集在水面,导致水华,随风浪飘动,受风浪的影响,主要聚集在离岸不远的水域。
2 成烃生物研究方法及识别
成烃生物的识别依传统常规的古生物研究方法,先将样品磨制成岩石光薄片,然后在透射光、反射光和荧光下,依据光学显微镜下见到的生物组织器官和叶状体形态进行成烃生物鉴定。而营底栖生活的海洋藻类主要包括红藻门红藻纲(除蓝色藻目外)、绿藻门石莼纲和异鞭藻门褐藻纲,其叶状体呈片状的薄壁组织和丝状集合体的假薄壁组织,与球状浮游藻类明显不同。另外,这些底栖藻类的生殖器官以及细胞结构也与浮游藻类也不同,如红藻多具有球状或带状三分和四分孢子囊,多数红藻类细胞间具有纹孔连接;褐藻具有单室或多室孢子囊,细胞壁之间存在细胞联丝和小孔[19-20]。因此,藻类鉴定主要是在光学显微镜下对藻类形态、组织器官以及细胞结构进行研究。
底栖藻类,一般具假根,根植于软质泥质基底,固定生长。保存在沉积岩中的底栖藻类多以叶状体残片为主,他们是鉴定底栖藻类的主体。底栖藻类叶状体具有一定的厚度,在保存条件好的情况下可见叶状体皮层和髓部分化清晰。如图2a所示,髓部细胞网状排列,皮层部分为含叶绿素细胞紧密排列,与髓部可明显分开。在塔里木盆地下古生界常见的底栖藻类大部分为红藻,红藻叶状体内部还发育孢子囊。若孢子囊脱出,在红藻叶状体出现凹窝(图2a),孢子囊及其内部孢子是红藻的生殖器官,为红藻囊果(图2b)。因此,显微镜下片状、块状的叶状体碎片以及球状囊果的出现是底栖藻类鉴定的重要标志。
而浮游藻类,无论是超微型浮游藻类还是大型浮游藻类,往往呈球状,在形态上与底栖藻类的片状区别明显。对于超微型浮游藻类而言,由于需要提高比表面积获得生态竞争优势,超微型浮游藻类往往呈单个球粒(图2c),而大型浮游藻类则一般呈集合的蚕豆状和球状(图2d)。
3 塔里木盆地早古生代成烃生物地球化学特征
不同的成烃生物除了具有生物特征差异外,例如特征的生物形态和组织结构等,这些特征可在显微镜下进行研究[1],还应有对石油地质学更为重要的有机地球化学和同位素地球化学特征差异,例如有机碳含量(TOC)和碳同位素值(δ13C)。这些地球化学特征是烃源岩评价和油-源对比的重要基础指标[13]。为建立起成烃生物与石油地质之间的桥梁,我们以塔里木盆地北缘4条下古生界剖面(南雅尔当山、苏盖特布拉克、东二沟和大湾沟剖面)(图3)为对象,对保存其中的沉积有机质进行显微镜下成烃生物辨识和常规地球化学特征研究。共采集新鲜样品66件,涉及层位包括寒武系西山布拉克组(1xs)、西大山组(1xd)、莫合尔山组(2m)、玉尔吐斯组(1y)、肖尔布拉克组(1x)和奥陶系吐尔沙克塔格组(O1tr)、黑土凹组(O1-2h)、萨尔干组(O2-3s)。
3.1 不同类型成烃生物烃源岩有机碳含量特征
在烃源岩评价中,有机碳含量(TOC)是一个非常直接而且有效的指标。然而,不同沉积环境、不同生态条件下形成的具有相同有机碳含量的烃源岩,即使在镜质体反射率(Ro)相似的条件下,其生烃能力还是存在一定的区别。已有的模拟实验也表明,不同生活习性的藻类,其生烃量和生烃高峰时期存在明显的差别[3]。因此,查明烃源岩中高TOC的生物贡献者将有利于对生烃潜力进行精细评价。
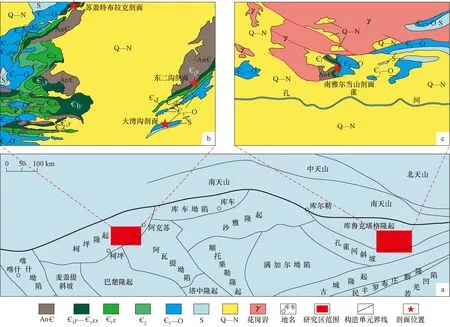
图3 研究区地质略图(a)及剖面位置(b,c)Fig.3 Simplified geological map showing the study area(a) and the locations of the sections(b,c)
如图4所示,在东二沟剖面玉尔吐斯组下段黑色泥页岩TOC存在明显的两分性,底部黑色泥页岩的TOC明显偏低,均在1%附近,而上部黑色泥页岩的TOC绝大部分超过10%。相应的成烃生物研究发现,底部低TOC段多为浮游藻类为主,含底栖藻类,而上部高TOC段多以底栖红藻为主,含浮游藻类,表明阿克苏地区玉尔吐斯组烃源岩的高TOC可能与底栖红藻存在一定的关联。这可能与不同生物有机质的成源和生烃过程的分解有关。在生烃过程中,等量底栖藻类的生烃能力只有浮游藻类的一半,意味着底栖藻类在生烃过后保留在地层中的残余TOC要明显高于浮游藻类。另外,在有机质氧化分解的过程中,底栖藻类抗氧化分解能力要强于浮游藻类,该过程导致了以底栖藻类为主的烃源岩的TOC要高于以浮游藻类为主的烃源岩。该认识在库鲁克塔格地区南雅尔当山剖面得到进一步印证,西山布拉克组下段以底栖藻类为主的样品的TOC要高于以浮游藻类为主的样品(图5)。
3.2 不同成烃生物组合烃源岩干酪根碳同位素特征
烃源岩碳同位素值(δ13C干酪根)是烃源岩的重要特征,也是进行烃源岩干酪根类型划分和油气源对比研究的重要指标。然而,目前关于不同成烃生物组合的烃源岩在干酪根碳同位素特征方面的研究较少。为此,本次研究对塔里木盆地下古生界烃源岩进行了干酪根碳同位素分析,结果见表1。
从表1可知,66件干酪根碳同位素值范围较大,在-38‰~-25‰。其中,南雅尔当山剖面西山布拉克组的δ13C干酪根为-34.2‰~-25.5‰,平均值为-30.3‰;西大山组δ13C干酪根在-32.8‰~-27.6‰,平均值为-31.1‰;莫合尔山组δ13C干酪根在-31.3‰~-30.0‰,平均值为-30.8‰;突尔沙克塔格组的δ13C干酪根在-32.4‰~-26.8‰;黑土凹组δ13C干酪根在-32.4‰~-26.8‰。苏盖特布拉克剖面玉尔吐斯组δ13C干酪根在-35.1‰~-26.5‰,平均值为-31.7‰;肖尔布拉克组δ13C干酪根在-32.2‰~-31.3‰。东二沟剖面玉尔吐斯组δ13C干酪根在-38.6‰~-28.3‰,平均值为-34.5‰。大弯沟剖面萨尔干组δ13C干酪根在-30.4‰~-29.2‰,平均值为-29.8‰。

图4 塔里木盆地北缘东二沟剖面玉尔吐斯组有机碳含量(TOC)与成烃生物组合Fig.4 TOCs and HOGs of the Yuertusi Formation in the Dongergou section,the north margin of the Tarim Basin(+表示数量较少;++表示数量中等;+++表示数量较多。)
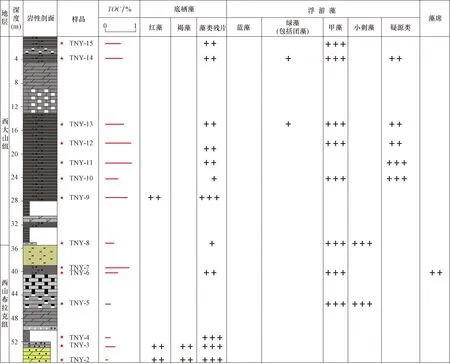
图5 塔里木盆地北缘南雅尔当山剖面西山布拉克组-西大山组有机碳含量(TOC)与成烃生物组合Fig.5 TOCs and HOGs of the Xishanbulake and Xidashan Formations in the Nanyaerdangshan section,the north margin of the Tarim Basin(+表示数量较少;++表示数量中等;+++表示数量较多。)
为进一步明确不同成烃生物烃源岩干酪根碳同位素组成的差异及规律,我们对4条剖面的干酪根碳同位素值与成烃生物组合进行了对比研究(图6)。结果发现,总体上以底栖藻为主的烃源岩,δ13C干酪根小于-34‰;以浮游藻为主的烃源岩,δ13C干酪根大于-30‰(表1;图6)。
在塔里木盆地东北缘库鲁克塔格地区的南雅尔当山剖面,西山布拉克组底部以底栖藻类为主,测得其δ13C干酪根均值为-34.2‰;以浮游藻类为主的西大山组和莫合尔山组,其δ13C干酪根均值为-30.2‰;而在线叶植物开始出现的突尔沙克塔格组和黑土凹组下段,δ13C干酪根均值变为-28.3‰;到黑土凹组上部以浮游藻为主,偶见底栖红藻,δ13C干酪根均值下降到-30.3‰(表1;图6)。
同样,在塔里木盆地西北缘柯坪地区苏盖特布拉克剖面玉尔吐斯组,以底栖藻类为主的样品TAS-1,TAS-11,TAS-13和TAS-15,δ13C干酪根范围为-35.1‰~-34.7‰;而以浮游藻类为主的样品TAS-2,TAS-9和TAS-16,δ13C干酪根范围为-28.5‰~-26.5‰。该剖面肖尔布拉克组下段以浮游藻类为主、含少量底栖藻类的3件样品,其δ13C干酪根为-32.6‰~-31.0‰。同样,在东二沟剖面玉尔吐斯组的研究发现,除TDE-8样品δ13C干酪根为-28.3‰(以绿藻、团藻、小刺藻和球状甲藻等浮游藻类为主)以外,其他以底栖藻类为主的样品δ13C干酪根普遍偏轻,在-38.6‰~-34.1‰。
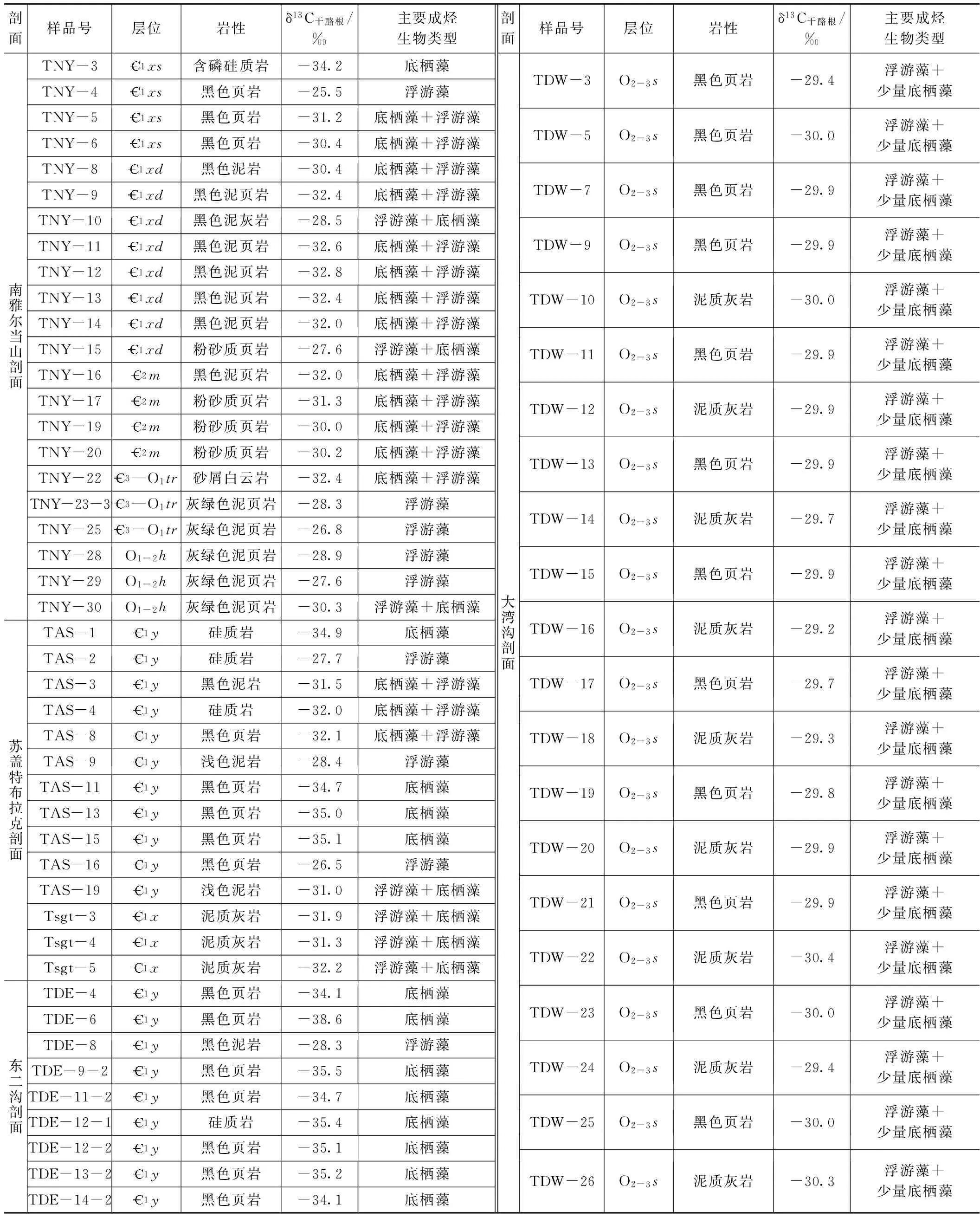
表1 塔里木盆地下古生界烃源岩干酪根碳同位素值及主要成烃生物类型
对大弯沟剖面奥陶系萨尔干组而言,成烃生物相对简单,成烃生物主要以蓝藻藻席和球藻等浮游藻类为主。与成烃生物镜下观察相匹配,δ13C干酪根范围非常窄,绝大多数在-30.4‰~-29.7‰(表1;图6)。
综上所述,以底栖藻类为主的烃源岩,δ13C干酪根小于-34‰;以浮游藻类为主的烃源岩,δ13C干酪根大于-30‰。
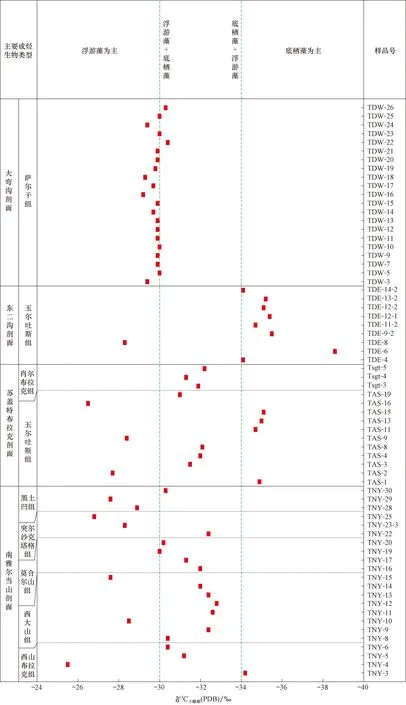
图6 塔里木盆地下古生界烃源岩干酪根碳同位素组成与成烃生物组合Fig.6 Carbon isotopic composition of kerogens and hydrocarbon generating organisms in the Lower Paleozoic source rocks of the Tarim Basin
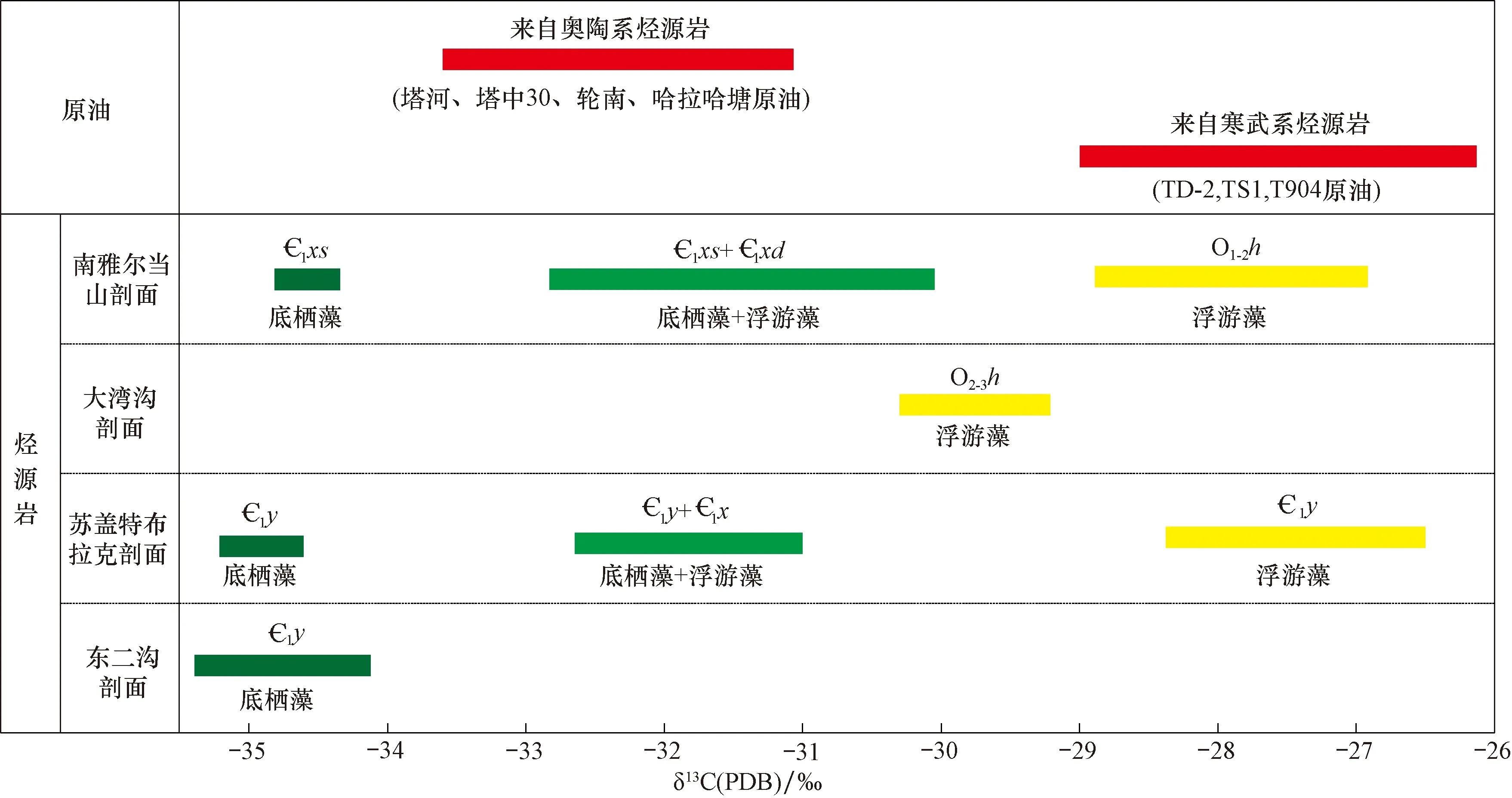
图7 塔里木盆地不同生物类型烃源岩干酪根碳同位素与原油碳同位素对比Fig.7 Comparison of carbon isotopes in kerogens of source rocks with different types of organisms and those in crude oil from the Tarim Basin
4 塔里木盆地古生界海相层系油气与成烃生物组合关系
根据统计前人分析资料和数据,塔里木盆地台盆区典型寒武系烃源岩和上奥陶统烃源岩在碳同位素组成上具有较明显的差别。图7对比了塔里木盆地寒武系与中、上奥陶统典型烃源岩全油及组分碳同位素组成。由图可见,除部分井寒武系原油碳同位素较重外,绝大部分原油碳同位素值分布在-34‰~-30‰;其中,-33.0‰~31.5‰的原油占70%。
依据油气生成过程中原油与干酪根之间碳同位素的继承关系,原油碳同位素比值一般要比其生油岩干酪根碳同位素比值小[13]。对比干酪根碳同位素值可见,寒武系原油比所有发现含底栖藻类的寒武系烃源岩的碳同位素都要轻,说明寒武系原油不可能来自寒武系底栖藻类烃源岩;但在苏盖特布拉克剖面发现的寒武系玉尔吐斯组中,浮游藻类的碳同位素值与寒武系原油基本相当,也可能作为寒武系原油的烃源岩(图7)。而对于塔河、塔中、轮南和哈拉哈塘油田原油而言,其原油碳同位素同时轻于寒武系和奥陶系的含浮游藻类烃源岩。从这个角度来说,不能断言奥陶系原油的烃源岩是哪个层位。综合寒武系、奥陶系原油与干酪根碳同位素对比研究,发现虽然不能确定烃源岩的层位,却可以断定底栖藻类对已发现原油的贡献不太可能,早古生界的浮游藻类是重要生烃母质。
5 结论
1) 藻类是早古生代最主要的成烃生物,按照生活习性,可以分为底栖藻类和浮游藻类。底栖藻类主要生活在海岸带,成带状分布,主要受光的强度、基底情况等因素控制;浮游藻类主要生活在海水表面,在不同海域的分布主要受藻体大小和风浪强度控制。
2) 受成源和生烃分解过程的影响,以底栖藻类为主的烃源岩的有机质碳含量高于以浮游藻类为主的烃源岩。
3) 对塔里木盆地北缘4条剖面的烃源岩干酪根碳同位素值与成烃生物组合面貌进行了对比研究,发现总体上以底栖藻类为主的烃源岩,δ13C干酪根小于-34‰;以浮游藻类为主的烃源岩,δ13C干酪根大于-30‰。对比塔里木盆地已发现原油碳同位素(均重于-30‰),初步推断其原油主要来源于浮游藻类的贡献。
致谢:感谢南京大学边立曾教授和中国石化石油勘探开发研究院黎茂稳教授在研究过程中提供的帮助,感谢北京大学张巍博士、中国石化石油勘探开发研究院无锡石油地质研究所罗厚勇博士、储层林博士和杨鑫博士在野外样品采集过程中提供的帮助。
[1] 边立曾.海相烃源岩生烃母质生物构成特征[C]∥关德范,秦建中,曹寅.石油地质样品分析测试技术及应用.北京:石油工业出版社,2006:269-290.
Bian Lizeng.Biological composition of oil generating material in marine source rocks[C]∥Guan Defan,Qin Jianzhong,Cao Yin.Application of analysis and testing technology in petroleum samples.Beijing:Petroleum Industry Press,2006:269-290.
[2] 谢树成,赖旭龙,黄咸雨,等.分子地层学的原理、方法及应用实例[J].地层学杂志,2007,31(3):209-221.
Xie Shucheng,Lai Xulong,Huang Xianyu,et al.Principles,methodo-logy and application of molecular stratigraphy[J].Journal of Stratigraphy,2007,31(3):209-221.
[3] 秦建中,陶国亮,腾格尔,等.南方海相优质页岩的成烃生物研究[J].石油实验地质,2010,32(3):262- 269.
Qing Jianzhong,Tao Guoliang,Tengger,et al.Hydrocarbon-forming organisms in excellent marine source rocks in south china[J].Petroleum Geology & Experiment,2010,32(3):262-269.
[4] 殷鸿福,谢树成,秦建中,等.对地球生物学、生物地质学和地球生物相的一些探讨[J].中国科学(D辑):地球科学,2008,38(12):1473-1480.
Yin Hongfu,Xie Shucheng,Qing Jianzhong,et al.Discussion on geobiology,biogeology and geobiofacies [J].Science in China (Series D:Earth Sciences),2008,38(12):1473-1480.
[5] 吴庆余,盛国英,傅家谟.一种蓝细菌的热解产气模拟[J].科学通报,1988,(18):1419-1421.
Wu Qingyu,Sheng Guoyin,Fu Jiamo.Thermal stimulation for cyanobacteria[J].Chinese Science Bulletin,1988,18:1419-1421.
[6] 张惠之,盛国英,王开发,等.盘星藻热模拟生油研究[J].同济大学学报(自然科学版),1994,23(3):291-299.
Zhang Huizhi,Sheng Guoying,Wang Kaifa,et al.Study on the thermal simulation for generating oil by pediastrum[J].Journal of Tongji University(Natural Science Edition),1994,23(3):291-299.
[7] 宋一涛,李树.颗石藻生烃的热模拟实验研究——(Ⅰ)烃的产率、性质及烯烃、烷烃的特征[J].高校地质学报,1995,1(2):95-106.
Song Yitao,Li Shu.Experimental study on simulating hydrocarbon generation of coccolithophoridaes by heating—(Ⅰ).generation rate and property of hydrocarbons,characteristic of alkanes and alkenes[J].Geological Journal of China Universities,1995,1(2):95-106.
[8] 李超,徐茂泉,王开发,等.单细胞海藻热模拟生烃研究[J].厦门大学学报(自然科学版),2001,40(3):764-769.
Li Chao,Xu Maoquan,Wang Kaifa,et al.Study on the thermal simulation for hydrocarbon generating by unicellular algae[J].Journal of Xiamen University (Natural Science Edition),2001,40(3):764-769.
[9] 郭汝泰,杨凤丽.藻类有机质的成烃机制探讨[J].同济大学学报(自然科学版),2002,30(1):41-45.
Guo Rutai,Yang Fengli.In question to the hydrocarbon generation mechanism for algal organic matter[J].Journal of Tongji University(Natural Science Edition),2002,30(1):41-45.
[10] 刘文斌,陆现彩,秦建中.海带热模拟实验的固体残余物研究及其石油地质意义[J].南京大学学报(自然科学版),2005,41(3):234-244.
Liu Wenbin,Lu Xiancai,Qin Jianzhong.Soil residue of thermal simulation experiment of laminaria and its petroleum geological significance[J].Journal of Nanjing University(Natural Sciences Edition),2005,41(3):234-244.
[11] 叶云,刘文汇,腾格尔,等.巢湖蓝藻腐殖化过程中形态与成分变化研究[J].微体古生物学报,2012,29(2):152-160.
Ye Yun,Liu Wenhui,Tengger,et al.Research on cyanobacteria from the chaohu lake during a simulating process of decaying:Changes in morphology and organic composition[J].Acta Micropalaeontologica Sinica,2012,29(2):152-160.
[12] 张水昌,梁狄刚,张宝民,等.塔里木盆地海相油气的生成[M].北京:石油工业出版社,2004.
Zhang Shuichang,Liang Digang,Zhang Baoming,et al.Marine hydrocarbon generating of Tarim[M].Beijing:Petroleum Industry Press,2004.
[13] Tissot B P,Welte D H.Petroleum formation and occurrence [M].Berlin,Germany:Springer-Verlag,1984:699.
[14] Brocks J J,Buick R,Summons R E,et al.A reconstruction of Ar-chean biological diversity based on molecular fossils from the 2.78~2.45 billion year old Mount Bruce Supergroup,Hamersley Basin,Western Australia[J].Geochim Cosmochim Acta,2003,67(22):4321-4335.
[15] Tappan H.The paleobiology of plant protists[M].San Francisco,USA:W H Freeman and Co,1980:1028.
[16] Butterfield N J.Bangiomorpha pubescensn gen,n sp:Implications for the evolution of sex,multicellularity,and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes[J].Paleobiology,2000,26 (3):386-404.
[17] Moldowan J M,Talyzina N M.Biogeochemical evidence for dinoagellate ancestors in the Early Cambrian[J].Science,1998,281:1168-1170.
[18] Stephen D K,Vanessa J K.An introduction to organic geochemistry[M].New Jersey,USA:Wiley Blackwell,2005:408.
[19] Fott B.Algenkunde [M].Jena,Germany:Veb Gustav Fischer Verlag,1971.
[20] Lee R E.Phycology [M].Cambridge,UK:Cambridge University Press,2008.
(编辑 李 军)
Organism assemblages in the Paleozoic source rocks and their implications
Liu Wenhui1,Hu Guang2,Tenger1,Wang Jie1,Lu Longfei1,Xie Xiaoming1
(1.WuxiBranchofPetroleumExplorationandProductionResearchInstitute,SINOPEC,Wuxi,Jiangsu214151,China; 2.SchoolofGeoscienceandTechnology,SouthwestPetroleumUniversity,Chengdu,Sichuan610500China)
The accumulation and preservation of the Paleozoic organisms provided a solid organic matter foundation for hydrocarbon generation during the Paleozoic.As different types of organisms were considered to contribute differently to hydrocarbon-generation,a study on organism assemblages (HGOs) started to prevail in source rock researches and has been used a new means of assessing the quality of matured source rocks.From geological,historical,and organic evolution perspectives,algae are the most important material providers for hydrocarbon generation during the early Paleozoic.They can be classified into two types according to their living habit: the benthic and the planktonic algae.The former lived near coast in a zonal distribution in accordance with light intensity and nutrient conditions.The latter lived in a photic zone and their distribution was controlled by their sizes and wind wave strength.Study on the HGOs and the organic geochemical characteristics of the source rock samples from seven formations of four Paleozoic outcrop sections shows that the TOCs in the source rocks dominated by benthic algae are higher than those controlled by planktonic algae.Together with depositional settings,the digenesis and hydrocarbon generation processes were all thought to have played some parts in shaping the organic carbon isotopic compositions of the samples.However,the processes seemed to have exerted lesser effect.Comparison of the kerogen carbon isotopic composition and the HGOs assemblages of the samples indicates that the δ13Ckerogenvalues of samples dominated with benthic algae are less than -34‰,whereas the δ13Ckerogenvalues of source rocks dominated with planktonic algae are more than -30‰.Taking the fact that the δ13C values (all more than -30‰) of known oil samples in the Tarim Basin into consideration,this article concludes that the crude in the basin is mostly sourced from rocks with planktonic algae and the source rocks with lower TOCs and planktonic algae from the Paleozoic may have greater exploration potential.
hydrocarbon generating organism,TOC,kerogen carbon isotope,oil-source correlation,Early Paleozoic,Tarim Basin
0253-9985(2016)05-0617-10
10.11743/ogg20160501
2016-04-06;
2016-09-13。
刘文汇(1957—),男,博士、教授,天然气地质学及油气地球化学。E-mail:whliu.syky@sinopec.com。
国家重点基础研究发展计划(973计划)项目(2012CB214801)。
TE122.1
A
