酸马奶和维利的研究进展
2016-11-11刘振民
王 豪,刘振民,章 慧
(1.光明乳业股份有限公司乳业研究院,乳业生物技术国家重点实验室,上海 200436;2.上海必诺检测技术服务有限公司,上海 200436)
酸马奶和维利的研究进展
王 豪1,刘振民1,章 慧2
(1.光明乳业股份有限公司乳业研究院,乳业生物技术国家重点实验室,上海 200436;2.上海必诺检测技术服务有限公司,上海 200436)
为迎合中国消费者日益增长的差异化饮食需求,本文通过综述传统发酵乳制品中酸马奶和维利的感官理化特性、微生物菌群、益生功能及生产工艺,以期能够将此类别具一格的发酵乳制品引入国内,并通过本土化改良进而实现规模化生产,从而打造出新兴的发酵乳细分市场来满足消费需求。
酸马奶;维利;发酵乳
乳是微生物最为常见的栖身场所,在适当的温度和时间条件下可引发乳的自然酸化。存在于大千世界中不同来源的微生物塑造出了多样化的发酵乳,发酵乳所具备的特征风味和质构与微生物息息相关[1]。基于微生物所形成的不同代谢产物,发酵乳发酵过程可被划分为以下三类[2]:乳酸菌发酵、酵母菌-乳酸菌发酵、霉菌-乳酸菌发酵。
大部分发酵乳的发酵过程为乳酸菌发酵。酵母菌-乳酸菌发酵的产品中除含有乳酸菌外,发酵过程中还会产生二氧化碳、乙醇和其他特征性风味物质,因此,此类产品也被定义为含酒精型乳制品。酵母菌-乳酸菌发酵的乳制品在东欧和中亚等地区十分流行,其中开菲尔(Kefir)和酸马奶(Koumiss)是传统意义上最为典型的酵母菌-乳酸菌发酵乳,而前者已在很多国家实现了工业化生产。
霉菌-乳酸菌发酵的乳制品起源于北欧地区,此类发酵乳制品质构普遍较为稠厚,乳固形物水平较常规酸奶更高,而酸感却比常规酸奶更弱。此外,霉菌在发酵过程中赋予这类产品独特的、差异化的口感及风味,备受北欧地区消费者的青睐。与酵母菌-乳酸菌发酵乳相比,霉菌-乳酸菌发酵乳的市场容量更小。
1 酸马奶
酸马奶是一种源自高加索地区,使用酵母菌搭配乳酸菌发酵的乳制品。自公元13世纪伊始,酸马奶从高加索地区逐渐传至中亚的蒙古和中国西北部。在马可波罗游记中,意大利人第一次将酸马奶的美名传播到了西方世界。如今,酸马奶常见于哈萨克斯坦、阿塞拜疆及土耳其等中亚国家,其中在哈萨克斯坦和吉尔吉斯斯坦尤为流行[3]。虽然酸马奶拥有悠久的食用历史和文化,但是关于这种发酵乳的研究仍较为有限。
1.1 酸马奶的感官及理化特征
原料乳品种的差异化赋予了酸马奶独特的感官特性及理化特征。传统的酸马奶使用马乳发酵,马乳的理化性质不同于牛乳,前者的pH值略高(约为7.0)、乳糖含量较高而蛋白质及脂肪含量则较低[2](表1)。
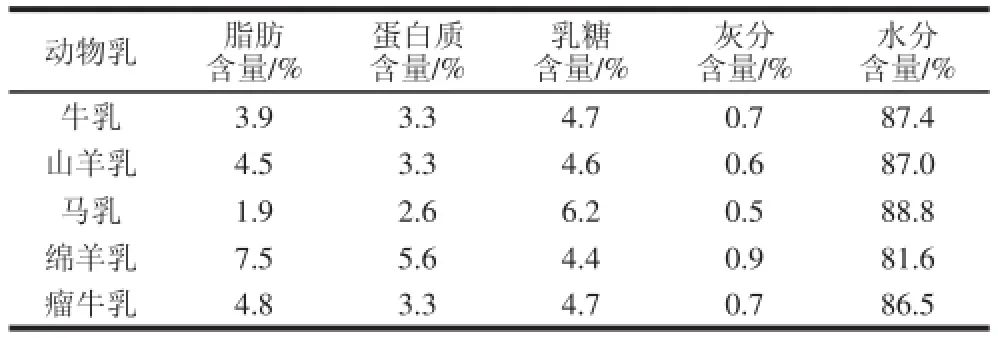
表1 不同动物乳中的组成成分[2]Table1 Chemical composition of milk of different animal species[[22]]
较高的乳糖含量使马乳呈透明的乳白色并略带浅灰色,口感较牛乳和山羊乳更甜。马乳中酪蛋白水平低于牛乳,但却有更多的免疫球蛋白[4]。通过对马乳中的蛋白质纯化后发现,马乳中的β-酪蛋白和α-酪蛋白较牛乳中相对应的蛋白质酸性更高[5]。此外,马乳中还含有高水平的多聚不饱和脂肪酸及磷脂[4,6]。基于马乳和牛乳的理化性质差异较为明显,发酵时牛乳的pH值下降幅度更大[7](图1)。
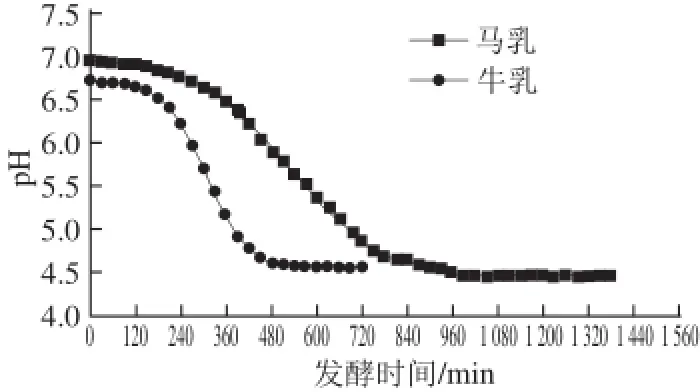
图1 马乳和牛乳的发酵酸化曲线[7]Fig.1 Acidification curves of mares’ milk and bovine milk during fermentation[7]
虽然多数发酵乳在发酵过程中会形成不同稠度的凝乳,但酸马奶在发酵结束后仍保持液体状态。这是由于马乳的蛋白质组成较为特殊,当酸度增加或添加凝乳酶时均不会形成凝乳。酸马奶这种独特的发酵特性使其在贮藏过程中乳清不易析出[1]。
酸马奶在发酵过程中,基于乳酸菌的同型发酵及异型发酵,兼之酵母菌的乙醇发酵,乳糖水平持续下降,而乳酸、乙醇、氨基酸和脂肪酸的含量升高。同时,发酵过程中还会产生多种风味物质(如乙酸、乙醇及酯类等),赋予产品清爽的发酵风味。表2所显示的即是酸马奶中营养成分的典型值[8]。

表2 酸马奶中营养成分的典型值[8]Table2 Typical values of nutrients in koumiss[8]
传统的酸马奶最高可含有3.0 g/100 mL的乙醇,且酸马奶中蛋白的水解程度(蛋白胨含量:0.2%~1.0%)较开菲尔(蛋白胨含量:0.05%~0.12%)高出近10 倍[9]。根据发酵程度的不同(乳酸及乙醇含量),Lozovich[9]将酸马奶划分为三大类型(表3)。Danova等[10]则根据酸马奶的pH值范围,亦将酸马奶划分为3 种不同风格:强烈(pH 3.3~3.6)、适中(pH 3.9~4.5)及轻柔(pH 4.5~5.0)。

表3 根据不同的发酵程度所划分的酸马奶[10]Table3 Classification of koumiss based on extent of fermentation[[1100]]
1.2 酸马奶中的微生物
酵母菌-乳酸菌发酵的产品中微生物复杂多变,这取决于发酵剂的来源地、微生物的生长条件、产品的加工过程及所使用的原料乳品种。一般而言,典型的酸马奶中菌群由乳酸菌、乳糖发酵型酵母菌和非乳糖发酵型酵母菌组成[11]。
酸马奶中较为常见的乳杆菌包括干酪乳杆菌、瑞士乳杆菌、植物乳杆菌、棒状乳杆菌、布氏乳杆菌、唾液乳杆菌、鼠李糖乳杆菌、副干酪乳杆菌副干酪亚种、副干酪乳杆菌坚忍亚种、马乳酒样乳杆菌、弯曲乳杆菌和发酵乳杆菌[10,12-13],较为常见的酵母菌包括马克斯克鲁维酵母、毕赤酵母和酿酒酵母[13-14]。
Robinson等[15]从蒙古酸马奶和中国酸马奶中不仅分离出乳杆菌(副干酪乳杆菌副干酪亚种、副干酪乳杆菌坚忍亚种及弯曲乳杆菌)和酵母菌(马克斯克鲁维酵母乳酸亚种和乳酒假丝酵母),还从蒙古酸马奶中分离出了乳球菌。由于乳球菌会加快发酵产酸速率,从而减慢酵母菌的增殖速率,最终导致酸马奶的特征风味减弱甚至丧失,因此在酸马奶的制备中需避免混入乳球菌。
Montanari等[16-17]收集调研了94 种来自哈萨克斯坦的传统酸马奶样品,发现主导乙醇发酵的优势菌群为单孢酵母(半乳糖发酵型酵母)。单孢酵母作为非乳糖发酵型酵母菌,会减慢乙醇发酵速率并产生多种代谢副产物(如甘油、琥珀酸、醋酸等),因此这类酸马奶常带有微弱的“异味”。
1.3 酸马奶的益生功能
酸马奶不仅是一种营养食品,更被牧民们看作是古老的治疗药物。酸马奶中微生物菌相与开菲尔相似,且同样包含诸如胞外多糖和活性肽等生物活性物质。在酸马奶流行的国度,民众普遍认为酸马奶是一种可以预防肠道紊乱、调节免疫平衡、辅助治疗肺结核等慢性疾病的益生食品。然而,与开菲尔相比,关于酸马奶益生方面的支持性科研数据极为有限。
酸马奶中的发酵菌群对许多病原菌(包括大肠杆菌、金黄色葡萄球菌、蜡样芽孢杆菌和结核分枝杆菌)具有拮抗或杀灭作用。在肠易激综合征康复期内,酸马奶可作为益生食品用于辅助治疗。Ham等[18]将从酸马奶中分离得到的乳酒假丝酵母混合植物乳杆菌喂食肉鸡后发现,粪便中的大肠杆菌数目明显下降。
血管紧张素Ⅰ-转化酶(angiotensin I-converting enzyme,ACE)的生物活性在血压调节中起着重要作用,ACE抑制剂具有显著的抗高血压作用。Sun Tiansong等[19]从酸马奶中分离出21 株乳杆菌,其中16 株表现出ACE抑制活性。Chen等[20]利用超滤技术及高效液相色谱(high performance liquid chromatography,HPLC)技术,从酸马奶中分离纯化出4 种具有ACE抑制活性的生物活性肽,序列分析显示其中1 种生物活性肽来源于马乳的β-酪蛋白。
di Cagno等[21]则发现,基于马乳中的过敏源物质水平低于牛乳,因此酸马奶更适用于过敏性儿童食用。
1.4 酸马奶的生产工艺
在早期,传统的酸马奶都是在马皮袋中经天然发酵制成的。通过多批次的发酵后(发酵结束后的成品从袋中排出后再灌入新批次的马乳),皮袋上残留的微生物菌群逐渐繁殖,从而成为传统酸马奶的发酵剂[15]。
现在,大部分蒙古人已改用木桶来替代马皮袋用于传统酸马奶的制作。蒙古人通常选择夏季(7-10月)来制作传统的酸马奶。牧民们在清晨时分开始挤奶,挤奶频率为2 h/次,每天平均挤奶6 次。待夜晚环境温度下降后,牧民们将鲜马乳灌至盛有残余酸马奶的木桶中,采用特制的器具剧烈搅拌1 h后,置于室温下持续发酵(图2),搅拌过程中混入的空气为酵母菌的增殖提供了良好的生长条件。马乳中的酵母菌数决定了终产品中的乙醇含量。根据蒙古人的制作经验,优质的酸马奶在发酵时所处的环境温度不宜过高。在蒙古,乙醇含量较高的酸马奶经蒸馏后还可获得馥郁芳香的蒸馏酒[11]。

图2 用于制备传统酸马奶的木制搅拌桨[11]Fig.2 Wooden mixing paddle used traditionally during the manufacture of koumiss[11]
1.4.2 改良方法
由于在当年的11月至次年的6月期间,母马的产奶量极为有限,因此人们试图以牛乳作为原料乳来生产“改良酸马奶”,从而使酸马奶摆脱季节性产品的束缚。基于马乳的蛋白质和乳糖含量不同于牛乳,牛乳中此类营养物的水平需进行模拟改良,才能生产出口味适宜的酸马奶。
较为经典的牛乳改性方法为:通过加水稀释脱脂牛乳来下调酪蛋白含量,补充乳清/乳清浓缩蛋白标准化至马乳中常规蛋白质水平,同时添加葡萄糖或乳糖(经β-D-半乳糖苷酶水解)标准化至马乳中常规碳水化合物水平[22]。
随着生产技术水平的提高,Kucukcetin等[23]成功地运用膜过滤技术(超滤、微滤及纳滤)对牛乳实施组分调整。发酵制成的产品经理化分析及感官评测证明,使用膜技术改进的牛乳可完全替代马乳用于生产酸马奶(表4、5)。在Kucukcetin等制备酸马奶的过程中,所使用的发酵菌种为马克斯克鲁维酵母乳酸亚种、德氏乳杆菌保加利亚亚种和嗜酸乳杆菌。
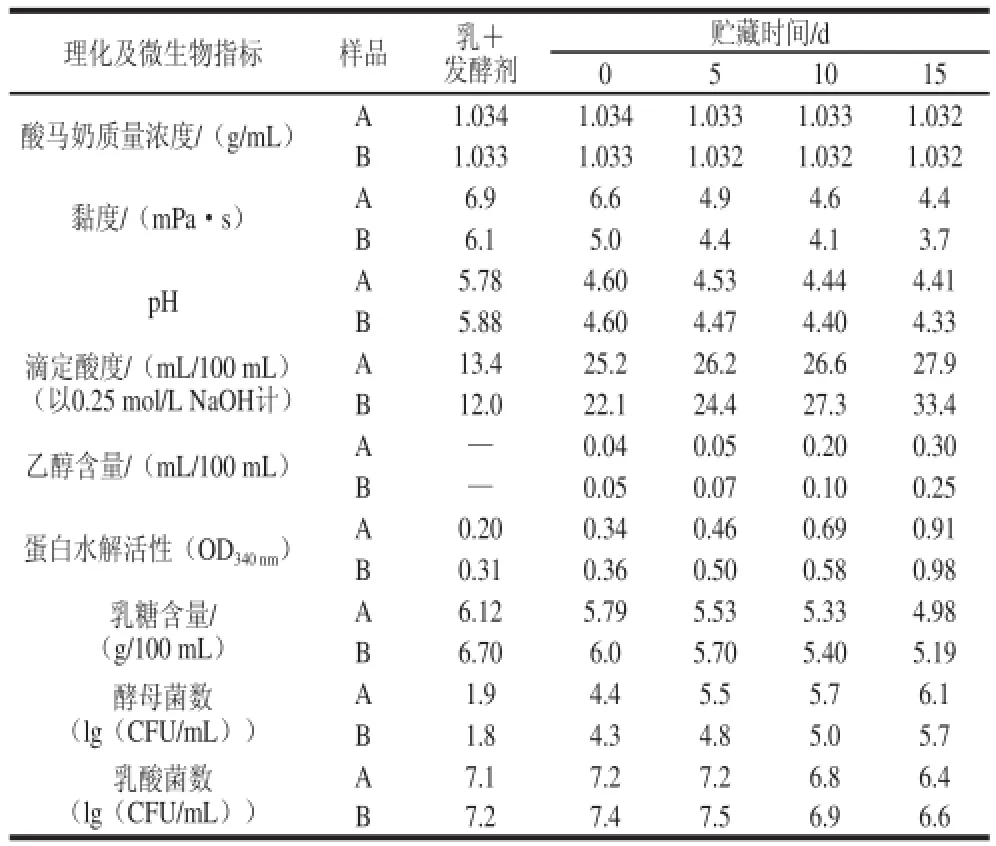
表4 使用改性牛乳或马乳制成的酸马奶在4 ℃贮藏期内的理化和微生物指标[23]Table4 Physical, chemical and microbiological properties of koumiss made from modified bovine milk or from mares’ milk during storage at 4 ℃[23]

表5 使用改性牛乳或马乳制成的酸马奶在4 ℃贮藏期内的感官指标[23]Table5 Organoleptic scores of koumiss made from modified bovine or from maarreess’ milk during storage at 4 ℃[23]
Yildiz等[2]记载了改良酸马奶的生产过程:改性牛乳经热处理后冷却至25 ℃,使用天然发酵剂或由分离自酸马奶的特征菌种组成的商业发酵剂进行发酵,发酵过程中不时地振荡搅拌,pH值为4.6时终止发酵,灌装封口后低温贮藏(图3)。与常规发酵乳不同之处在于,在生产过程中,基于所采用的原料乳在酪蛋白等电点无法形成凝乳的特性,酸马奶不适于制作成凝固型产品。
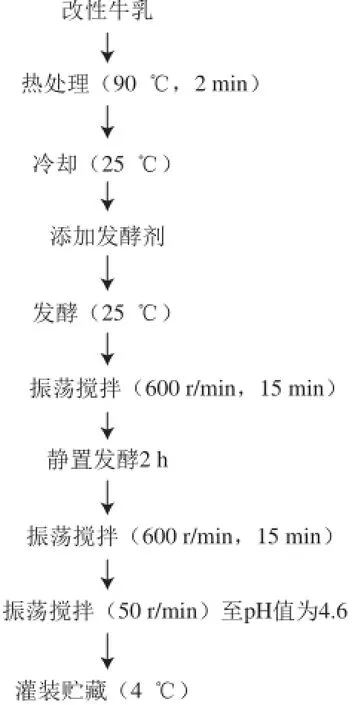
图3 改良酸马奶的生产工艺[2]Fig.3 Production scheme for making modified koumiss[2]
与开菲尔的相似之处在于,酸马奶同样会因酵母菌产气而引发涨包现象。当打开玻璃瓶时,可能有多达1/3的内容物会产生大量泡沫,如果产品的包装为纸盒,包装容器可能还会发生爆炸。为避免上述问题的发生,可选择在灌装前向包装容器内充填N2,或选择在灌装后使用N2取代包装容器的顶空部分[22]。
[4] Adam Entous, “In Asia, Tone Lightens On Sea Disputes,” Wall Street Journal, October 13, 2010.
2 维利2
维利(viili)是由传统产品菲慕克(Filmjölk)衍变过来的现代产品。在芬兰,该产品的人均年消费量为4.5 kg/(人·a),是最受欢迎的发酵乳饮料。如今维利已在多个北欧国家上市,包括丹麦、瑞典、挪威和冰岛。在北欧国度的各大卖场甚至是小卖铺,均可买到美味的维利。
维利的类型多种多样,除传统的原味和水果味外,市场上也可以见到低脂、低乳糖、低热量和添加益生菌等具有特殊功能诉求的产品系列。北欧地区的居民习惯在早餐时候享用维利,在他们的生活中维利是一种常见的甜点。随着现代化节奏的加快,维利已开始转型为快餐食品。
2.1 维利的感官特征
传统的维利表面是一层白色的霉菌,赋予产品天鹅绒般柔软的外观。此霉菌可分解代谢凝乳中的乳酸,从而降低产品的酸度。因此,维利的口感柔和、酸甜度适中且具有独特的发酵风味[24],其风味与酪乳较为类似。此外,霉菌通过好氧发酵产生的二氧化碳还使产品具有轻微的含气感。在食用维利时,人们经常还会添加肉桂粉等香料来赋予其特殊的香气。
维利的质地均一、黏稠度较高,具有轻度的拉丝性。凝乳很容易用勺子挖取,因此可被进一步加工成具有多种风味的新鲜奶酪。通常,维利用汤勺食用,如此凝胶状的产品会很容易切成小块。如果将维利搅拌混匀后食用,产品的拉丝性会变得非常明显。
维利中含有大量的胞外多糖,受益于菌种发酵所产生的这种多糖,产品的颗粒感较弱且脱水收缩的现象较少[25]。当从容器中挖取一勺维利,放置于盘子上,这勺维利会保持原型,不至于塌陷或者失去硬度,并且在2 min内不会出现乳清析出(图4)。

图4 维利的凝乳及凝乳切块[11]Fig.4 Curd and slices of viili[11]
2.2 维利中的微生物
维利发酵剂中常见的乳球菌分为产黏型的乳酸乳球菌乳酸亚种和非产黏型的乳酸乳球菌乳脂亚种[26]。产黏型乳球菌和非产黏型乳球菌分别单独发酵时体系的微观结构差异较大(图5、6)[27]。许多产黏型乳球菌的产酸能力较弱,因此在发酵剂中常搭配产酸较快的非产黏型乳球菌以提升发酵速率。此外,在维利的优势菌群中还经常发现乳酸乳球菌乳亚种双乙酰变种和肠膜明串珠菌乳脂亚种,这类嗜温型乳酸菌可产生多种芳香类物质。典型的维利中还含有白地霉,白地霉是一种兼具酵母菌和霉菌特性的丝状真菌,在严格好氧条件下此霉菌会形成密集、柔软的绒毛层(类似于卡门培尔干酪和布里干酪),覆盖整个产品表面。

图5 分离自维利的非产黏型乳球菌发酵乳的扫描电子显微镜图[27]Fig.5 Scanning electron micrograph of fermented milk made with non-ropy Lactococcus isolated from viili[27]
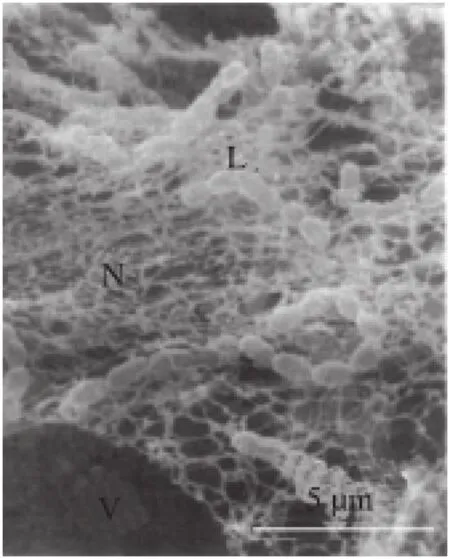
图6 分离自维利的产黏型乳球菌发酵乳的扫描电子显微镜图[27]Fig.6 Scanning electron micrograph of fermented milk made with ropy Lactococcus isolated from viili[27]
在维利的发酵剂生产过程中,噬菌体污染事件时有发生。噬菌体会抑制菌体细胞的增殖并延长产品的发酵时间。因此,针对噬菌体的专一性,菌种生产商们往往使用轮换菌种来避免噬菌体污染现象的发生。然而,目前仅有极少数的产黏型乳球菌拥有可替代的轮换菌种。
产黏型乳球菌在代谢过程中可分泌大量胞外多糖,胞外多糖不仅表现出特殊的流变学特性,同时还能保护菌体细胞免受噬菌体的攻击[26],并作为益生元促进肠道内益生菌群的增殖[28]。然而,乳球菌产胞外多糖的生物功能稳定性较差,重复传代的次数增多或培养温度出现异常均会弱化菌种的胞外多糖产生能力,甚至导致菌体细胞无法合成胞外多糖。Vedamuthu等[29]发现产胞外多糖的相关基因簇位于细胞的质粒中,进而通过质粒接合转移技术将非产黏型乳球菌成功转化为产黏型乳球菌,相关结果最终得到了专利授权。van Kranenburg等[30]进一步证实了乳球菌的异型胞外多糖产生受一个43 kb质粒的控制,该质粒包含一个12 kb胞外多糖基因组。
2.3 维利的益生功能
维利作为北欧传统发酵乳的典型代表,具有一定的免疫调节作用,主要源于发酵乳中的乳球菌表面含有抗原结构。乳酸乳球菌乳脂亚种是维利中的主发酵剂,据报道该菌种不仅能刺激机体分泌免疫球蛋白IgM,同时还能促进T淋巴细胞的增殖[31]。此外,该乳球菌还可激活S180肉瘤细胞小鼠腹腔内巨噬细胞的吞噬活性。通过腹腔注射该菌种的冻干制剂后,可延缓小鼠腹水瘤及实体瘤的生长。经沙门氏菌回复突变实验(亦称Ames实验)验证,乳酸乳球菌乳脂亚种可降低亚硝胺的致突变作用,降幅达40%[1]。Kitazawa等[31]则研究表明,分离自维利的乳球菌除能抑制金黄色葡萄球菌和大肠杆菌外,亦可降低小鼠的血清胆固醇水平。
2.4 维利的生产工艺
维利的工业化生产起于20世纪50年代晚期。生产过程中,标准化的原料乳(乳脂含量介于2%~12%之间)均质或不经均质,加热至82~83 ℃后保温20~25 min或经改良的高温短时(high temperature short time,HTST)杀菌(78 ℃,2 min)后,冷却至20 ℃并添加发酵剂。发酵剂由嗜温型乳酸菌(乳酸乳球菌乳酸亚种、乳酸乳球菌乳脂亚种、乳酸乳球菌乳亚种双乙酰变种和肠膜明串珠菌乳脂亚种等)和白地霉组成。灌装后在20 ℃条件下发酵20~24 h至滴定酸度达0.9%(或终点pH值达4.6)。随后产品冷却至6 ℃以下贮藏[1]。采用此法生产的维利拥有至少14 d的货架期。
未经均质的维利顶部会形成较厚的脂肪层,可将霉菌充分包裹并赋予产品典型的霉菌风味[24]。然而,霉菌的生长易受包装容器顶部氧气量的影响。如果盖子破损或者密封不当,霉菌会继续生长,继而形成浓稠且不规则的表层,致使产品产生不可接受的霉味。
3 结 语
酸马奶与常规发酵乳的最大区别在于,前者采用酵母菌协同乳酸菌发酵马乳,同时发酵过程中通过产生二氧化碳和乙醇,赋予产品独特的杀口感和酒香味。然而,受限于较为刺激的口感,以及工业化生产过程中所存在的交叉污染现象,酸马奶的消费市场仍局限于传统地区,还未实现全球化的上市销售。
维利采用白地霉进行协同发酵,具有特殊的感官特征及发酵菌种,是所有发酵乳品类中差异化要素最显著的一类产品,一定程度上拓宽了生产者对发酵菌种的认知视野。然而,由于霉菌的发酵风味在非传统地区较难被当地消费者所接受,同时霉菌的使用在工业化生产过程中存在较高的质量安全风险,因此维利的销售区域极为有限。
酸马奶和维利所具有的独特感官特性及营养益生功能可满足当下消费者日益增长的高品质差异化需求,对现有同质化越发严重的发酵乳市场可形成较强的冲击力。然而,酸马奶和维利若想在未来取得长足进步,有必要针对乳制品消费密度较高的地区进行本土化改良,同时需降低大规模、多生产线操作模式下的污染风险,如此方可实现传统地区的酵母菌-乳酸菌发酵乳和霉菌-乳酸菌发酵乳在非传统地区的成功演绎。
[1] CHANDAN R C, KILARA A. Manufacturing yogurt and fermented milks[M]. 2nd ed. Chichester: John Wiley & Sons Inc., 2013: 6-18; 394-409; 447-448. DOI:10.1002/9781118481301.
[2] YILDIZ F. Development and manufacture of yogurt and functional dairy products[M]. Boca Raton: CRC Press, 2010: 144-159. DOI:10.1201/9781420082081.
[3] WANG J G, CHEN X, LIU W J, et al. Identification of Lactobacillus from koumiss by conventional and molecular methods[J]. European Food Research and Technology, 2008, 227(5): 1555-1561. DOI:10.1007/s00217-008-0880-4.
[4] MALACARNE M, MARTUZZI F, SUMMER A, et al. Protein and fat composition of mare’s milk: some nutritional remarks with reference to human and cow’s milk[J]. International Dairy Journal, 2002, 12(11): 869-877. DOI:10.1016/S0958-6946(02)00120-6.
[5] EGITO A S, MICLO L, LOPEZ C, et al. Separation and characterization of mare’s milk αs1-, β-, κ-caseins, γ-casein-like, and protease peptone comp onent 5-like peptides[J]. Journal of Dairy Science, 2002, 85(4): 697-706. DOI:10.3168/jds.S0022-0302(02)74126-X.
[6] IAMETTI S, TEDESCHI G, OUNGRE E. Primary structure of kappacasein isolated from mare’s milk[J]. Journal of Dairy Research, 2001, 68(1): 53-61. DOI:10.1017/S0022029900004544.
[7] BORNAZ S, GUIZANI N, SAMMARI J, et al. Physicochemical properties of fermented Arabian mares’milk[J]. International Dairy Journal, 2010, 20(7): 500-505. DOI:10.1016/j.idairyj.2010.02.001.
[8] LIU S N, YE H, ZHOU Z J. Lactic acid bacteria in traditional fermented Chinese foods[J]. Food Research International, 2011, 44(3): 643-651. DOI:10.1016/j.foodres.2010.12.034.
[9] LOZOVICH S. Medical uses of whole fermented mare milk in Russia[J]. Cultured Dairy Products Journal, 1995, 30(1): 18-20.
[10] DANOVA S, PETROV K, PAVLOV P, et al. Isolation and characterization of Lactobacillus strains involved in koumiss fermentation[J]. International Journal of Dairy Technology, 2005, 58(2): 100-105. DOI:10.1111/j.1471-0307.2005.00194.x.
[11] TAMIME A Y, ROBINSON R K. Fermented milks[M]. Oxford: Blackwell Science Ltd., 2006: 175-205. DOI:10.1002/9780470995501.
[12] ISHII S. Study on the koumiss (airag) of Mongolian nomads after severe cold winters of 2000 and 2001[J]. Milk Science, 2003, 52(1): 49-52.
[13] WATANABE K, FUJIMOTO J, SASAMOTO M, et al. Diversity of lactic acid bacteria and yeasts in airag and tarag, traditional fermented milk products from Mongolia[J]. World Journal of Microbiology and Biotechnology, 2008, 24(8): 1313-1325. DOI: 10.1007/s11274-007-9604-3.
[14] 倪慧娟, 包秋华, 孙天松, 等. 新疆地区酸马奶中酵母菌的鉴定及其生物多样性分析[J]. 微生物学报, 2007, 47(4): 578-582. DOI:10.3321/j.issn:0001-6209.2007.04.003.
[15] ROBINSON R K. Dairy microbiology handbook[M]. 3rd ed. New York: John Wiley & Sons, 2002: 367-490. DOI:10.1002/0471723959.
[16] MONTANARI G, GRAZIA L. Galactose-fermenting yeasts as fermentation microorganisms in traditional koumiss[J]. Food Technology and Biotechnology, 1997, 35(3): 305-308.
[17] MONTANARI G, ZAMBONELLI C, GRAZIA L, et al. Saccharomyces unisporus as the principal alcoholic fermentation microorganism of traditional koumiss[J]. Journal of Dairy Research, 1996, 63(2): 327-331. DOI:10.1017/S0022029900031836.
[18] HAM J S, JEONG J Y, JEONG S G, et al. Effects of feeding Lactobacillus plantarum and Candida kefyr isolated from Mongolian koumiss on the growth and fecal microflora of broilers[J]. Korean Journal of Dairy Science, 1999, 21(2): 241-246.
[19] SUN T S, ZHAO S P, WANG H K, et al. ACE-inhibitory activity and gamma-aminobutyric acid content of fermented skim milks by Lactobacillus helveticus isolated from Xinjiang koumiss in China[J]. European Food Research and Technology, 2009, 228(4): 607-612. DOI:10.1007/s00217-008-0969-9.
[20] CHEN Y, WANG Z, CHEN X, et al. Identification of angiotensin I-converting enzyme inhibitory peptides from koumiss, a traditional fermented mare’s milk[J]. Journal of Dairy Science, 2010, 93(3): 884-892. DOI:10.3168/jds.2009-2672.
[21] di CAGNO R, TAMBORRINO A, GALLO G, et al. Uses of mares’milk in manufacture of fermented milks[J]. International Dairy Journal, 2004, 14(9): 767-775. DOI:10.1016/j.idairyj.2004.02.005.
[22] SEILER H. A review: yeasts inkefir and koumiss[J]. Milchwissenschaft, 2003, 58(7/8): 392-396.
[23] KUCUKCETIN A, YAYGIN H, HINRICHS J, et al. Adaptation of bovine milk towards mares’ milk composition by means of membrane technology for koumiss manufacture[J]. International Dairy Journal, 2003, 13(12): 945-951. DOI:10.1016/S0958-6946(03)00143-2.
[24] KAHALA M, MAKI M, LEHTOVAARA A, et al. Characterization of starter lactic acid bacteria from the Finnish fermented milk product viili[J]. Journal of Applied Microbiology, 2008, 105(6): 1929-1938. DOI:10.1111/j.1365-2672.2008.03952.x.
[25] MACURA D, TOWNSLEY P M. Scandinavian ropy milkidentification and characterization of endogenous ropy lactic streptococci and their extracellular excretion[J]. Journal of Dairy Science, 1984, 67(4): 735-744. DOI:10.3168/jds.S0022-0302(84)81363-6.
[26] DEVEAU H, van CALSTEREN M R, MOINEAU S. Effect of exopolysaccharides on phage-host interactions in Lactococcus lactis[J]. Applied Environmental Microbiology, 2002, 68(9): 4364-4369. DOI:10.1128/AEM.68.9.4364-4369.2002.
[27] TOBA T, NAKAJIMA H, TOBITANI A, et al. Scanning electron microscopic and texture studies on characteristic consistency of Nordic ropy sour milk[J]. International Journal of Food Microbiology, 1990, 11(3/4): 313-320. DOI:10.1016/0168-1605(90)90024-Y.
[28] RUAS-MADIEDO P, GUEIMONDE M, MARGOLLES A, et al. Short communication: effect of exopolysaccharide isolated from “viili”on the adhesion of probiotics and pathogens to intestinal mucus[J]. Journal of Dairy Science, 2006, 89(7): 2355-2358. DOI:10.3168/jds. S0022-0302(06)72307-4.
[29] VEDAMUTHU E R, NEVILLE J M. Involvement of a plasmid in production of ropiness (mucoidness) in milk cultures by Streptococcus cremoris MS[J]. Applied Environmental Microbiology, 1986, 51(4): 677-682.
[30] van KRANENBURG R, BOELS I C, KLEEREBEZEM M. Genetics and engineering of microbial exopolysaccharides for food: approaches for the production of existing and novel polysaccharides[J]. Current Opinion in Biotechnology, 1999, 10(5): 498-504. DOI:10.1016/S0958-1669(99)00017-8.
[31] KITAZAWA H, TOBA T, ITOH T, et al. Antitumoral activity of slime-forming, encapsulated Lactococcus lactis subsp. cremoris isolated from Scandinavian ropy sour milk ‘viili’[J]. Animal Science and Technology, 1991, 62(3): 277-283. DOI:10.2508/chikusan.62.277.
Advances in Koumiss and Viili
WANG Hao1, LIU Zhenmin1, ZHANG Hui2
(1. State Key Laboratory of Dairy Biotechnology, Dairy Research Institute, Bright Dairy and Food Co. Ltd., Shanghai 200436, China; 2. Shanghai Bino Testing Service Co. Ltd., Shanghai 200436, China)
The sensory profiles, physicochemical properties, microflora, health benefits and manufacturing processes of koumiss and viili, as distinctive representatives of traditional fermented milk, have been summarized in this article. The introduction and large-scale production of koumiss and viili improved to meet the increasing requirements of Chinese consumers is beneficial for developing the emerging niche market in the field of fermented milk and further catering to changing consumer demands.
koumiss; viili; fermented milk
10.7506/spkx1002-6630-201603043
TS252.54
A
1002-6630(2016)03-0247-06
王豪, 刘振民, 章慧. 酸马奶和维利的研究进展[J]. 食品科学, 2016, 37(3): 247-252. DOI:10.7506/spkx1002-6630-201603043. http://www.spkx.net.cn
WANG Hao, LIU Zhenmin, ZHANG Hui. Advances in koumiss and viili[J]. Food Science, 2016, 37(3): 247-252. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201603043. http://www.spkx.net.cn
2015-05-28
“十二五”国家科技支撑计划项目(2012BAD28B07)
王豪(1984—),男,工程师,硕士,研究方向为乳品科学与技术。E-mail:wanghao840818@126.com
