冷冻-复温过程中葡萄相变过程研究
2016-10-27郝俊杰王怀文计宏伟
张 哲 郝俊杰 赵 静 王怀文 计宏伟 张 平
(1.天津商业大学天津市制冷技术重点实验室, 天津 300134;2.国家农产品保鲜工程技术研究中心天津市农产品采后生理与贮藏保鲜重点实验室, 天津 300384)
冷冻-复温过程中葡萄相变过程研究
张哲1郝俊杰1赵静1王怀文1计宏伟1张平2
(1.天津商业大学天津市制冷技术重点实验室, 天津 300134;2.国家农产品保鲜工程技术研究中心天津市农产品采后生理与贮藏保鲜重点实验室, 天津 300384)
为研究葡萄在冷冻-复温过程的相变过程变化,利用差示扫描量热仪DSC系统对葡萄果肉进行冷冻-复温实验。通过改变葡萄试样冷冻-复温过程的速率、次数、时间间隔、速率梯度以及通过最大冰晶生成带的速率等实验条件,分析实验过程中的热流曲线,对比冷冻-复温过程中的相变潜热、相变结束点、相变开始点以及相变峰值等参数变化,得到不同条件下冷冻-复温过程对葡萄试样的影响规律。研究发现:冷冻过程中速率越大对应的结冰点温度越低,时间越短。增加复温速率梯度能有效提高冰晶融化相变过程的潜热、峰点温度和结束点温度。复温速率由2℃/min变化到20℃/min过程中相变潜热增加了1.79倍,相变峰点温度增加了0.94倍,相变结束点温度增加了5.07倍。该研究对果蔬冷冻-复温过程后的品质恢复提供了依据。
葡萄果肉; 冷冻-复温; 相变; DSC系统
引言
随着生活水平的提高,人们对果蔬冷冻保鲜的要求越来越高,但是在贮运过程中,果蔬不可避免地进行着冷冻-复温过程,因此了解冷冻-复温过程中果蔬的相变过程变得尤为重要。在国外,THIRUMALA等[1]利用差示扫描量热仪DSC(Differential scanning calorimetry)分析了冻结过程中不规则形状细胞的体积变化规律,MORI等[2]使用DSC研究了细胞悬浮液冻结过程中的水分运输和胞内冰形成机理。DEVIREDDY等[3]用DSC分析测定了细胞膜的渗透性质。SORGENTINI[4]利用DSC研究了大豆分离蛋白和乳清蛋白的热属性,并通过峰温和吸热曲线所围成的面积计算对应的变性热焓。在国内,张素文[5]利用DSC研究了不同解冻方法对西兰花部分玻璃态贮藏品质的影响。樊振江等[6]利用DSC研究了速冻温度和速冻前漂烫过程对猕猴桃果丁的细胞结构和品质的影响。张哲等[7]利用低温显微镜系统研究了葡萄细胞在冷冻-复温过程的结晶变化,但只研究了冷冻-复温过程的葡萄细胞显微图像、细胞体积、内压、渗透率的变化,未采用DSC研究相变过程。
现有文献对果蔬在冷冻-复温过程中相变过程研究较少,本文以红提葡萄为实验对象,从葡萄潜热变化角度出发利用DSC系统对葡萄冷冻-复温过程中的相变过程进行研究,旨为果蔬冷冻-复温过程后的品质恢复提供依据。
1 实验材料与方法
1.1实验设备

图1 DSC系统Fig.1 DSC system
为研究冷冻-复温过程中果蔬物性的变化,实验采用美国TA 公司生产的Q1000差示扫描量热仪系统(图1),包括差示扫描量热仪、计算机数据采集系统和用于吹扫试样的高纯氮气(99.99%),实验环境可从-180℃变化到725℃。其工作原理是通过固体封压机将试样和参考样分别压装在坩埚内,设定实验参数,通过调节试样盘和参考样盘下面放置的加热片来调节试样和参考样的加热温度,并使其均匀受热。降温操作是利用高纯氮气对试样和参考样进行吹扫,流量控制在50 mL/min,以保证试样受冷均匀,实验过程中测得的数据直接记录在数据库中。
本实验所用的实验水果是新鲜红提葡萄,采购于天津市咸阳北路菜市场,统一选取果粒为卵圆形、松紧度适中的红提葡萄。
1.2实验方法
实验使用的坩埚为铝制固体坩埚,容积大约为50 mm3,先用电子天平称取坩埚质量,然后用镊子向坩埚中加入一定量待测试样,再次称量后用DSC专用压模固体封压机将坩埚压紧,放入DSC炉内,在计算机中设定好实验条件和参数,开始实验:
(1)首先挑选2套干净的坩埚,用电子天平分别称量,记录并编号,选择一个坩埚作为参考样,另一个准备盛放样品。
(2)选取少量葡萄果肉用镊子夹入盛放样品的坩埚内,再次用电子天平称量。
(3)分别用DSC专用压模器封好坩埚。
(4)把坩埚依次放入炉腔内。
(5)在计算机上设定相应程序:炉子温度在28℃维持3 min,消除环境因素的影响,设定不同的实验参数对试样进行冷冻-复温,保证高纯氮气的流速在50 mL/min,实验数据保存后,可由DSC自带分析软件中得到样品在冷冻-复温过程中的热流曲线。
1.3理论计算方法

图2 冷冻-复温过程的热流曲线Fig.2 Heat flow curve of freezing-thawing process
以某一速率下的冷冻-复温过程热流曲线为例,如图2所示。从图中可以看出相变过程释放的相变潜热远远高于无相变时的显热,因此,在DSC热流曲线上的相变过程会产生一个峰值。图中DSC的热流曲线向上方向的峰为放热峰,代表降温条件下的相变即结晶过程。向下方向的峰为吸热峰,代表升温条件下的相变过程即冰晶融化过程。吸热峰的相变过程初始点对应的温度为融化点温度,相变峰结束点为吸热过程结束点。相变峰的峰点代表最大的转换速率点,相变峰的面积代表相变过程潜热。在冷冻过程中,葡萄的温度会降低,当温度降低到过冷点b时开始发生相变。b点的温度为过冷温度,冻结过程释放的相变潜热使葡萄温度迅速升高到冰点a,葡萄全部结冰后温度继续降低,在复温过程中,c点为相变过程初始点即融化点,d点为最大的转换速率点即相变峰点,e点为相变过程结束点。这与曾彦彰[8]在预处理生物材料的差示扫描量热学研究中的结论一致。
由以上分析可知过冷度表达式为
ΔT=Ta-Tb
(1)
式中Ta、Tb——a、b点的温度
在DSC热分析图中对结晶的焓值进行积分可以得到相变潜热,计算式为
(2)
式中dT——温度无限小区域
dH——温度无限小区域(dT)中结晶的焓值
利用TA 公司自带软件TA Universal Analysis来分析热流曲线图,通过读取实验中热流曲线图上的相变峰面积可以直接求出相变过程的潜热变化,首先选择不同的基线类型,然后对热流曲线相变峰的面积进行积分运算,求出相变过程的潜热值,热流曲线的相变峰的面积积分公式为
(3)
式中S——相变峰面积
t1、t2——相变过程的开始点、结束点时间
ΔPC——相变过程前后热流曲线信号的差值
2 实验结果与分析
2.1冷冻过程的分析计算
在实验前用DSC对去离子水进行测定,测得融化点温度为0℃,与标准融化点0℃相同,潜热为334.9 kJ/kg,和标准相变潜热335 kJ/kg的误差小于0.1%,因此该仪器是可靠的[9-10]。用电子天平分别称量20 mg的葡萄果肉压装在坩埚内,利用DSC从室温28℃左右快速降温至-40℃,设定降温速率分别为2、6、8、11、14、20℃/min。热流曲线图中的相变潜热和结冰点温度变化以6、14℃/min为例,如图3所示。

图3 冷冻过程的热流曲线Fig.3 Heat flow curve of freezing process
用TA Universal Analysis软件选取线性基线,分析计算相变峰面积以求得葡萄在冷冻过程的相变潜热,得到葡萄相变过程的潜热和结冰点温度,如表1所示。
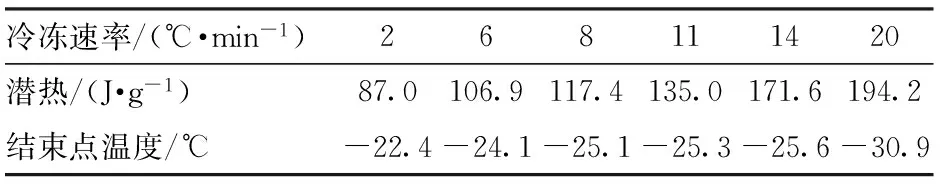
表1 冷冻过程中潜热和结冰点变化Tab.1 Change of latent heat and freezing point in the process of freezing

图4 相变潜热变化曲线Fig.4 Phase latent heat changes of grape

图5 结冰点温度变化曲线Fig.5 Freezing temperature changes of grape
由图4、5可知葡萄试样在冷冻过程中的相变潜热随降温速率的增大而增大,在2℃/min时相变潜热为87 J/g,在20℃/min时相变潜热为194.2 J/g,因此在降温速率由2℃/min到20℃/min变化过程中,相变潜热增大了1.23倍。而葡萄试样的冰点随降温速率的增大而降低,由2℃/min速率的-22.4℃变化到20℃/min速率的-30.9℃,冰点降低了37.9%。
冷冻速率越大,葡萄果肉的过冷度越大,在冷冻过程中释放的相变潜热越大,葡萄试样完成相变的时间越短,形成的冰晶不容易长大,因此会形成细小的冰晶,而降温速率越小,葡萄果肉的过冷度越小,完成相变的时间越长,形成的冰晶得以长大,因此会形成较大冰晶。细小的冰晶对葡萄细胞的机械损伤要小于较大冰晶,因此在冷冻过程中应适当加大冷冻速率。
2.2重复1次冷冻过程(冷冻后再解冻)
冷冻过程中,为探究不同速率条件下重复1次冷冻-复温过程对葡萄试样品质物性的影响,对葡萄试样先分别以2、6、8、11、14、20℃/min的速率降温到-40℃左右,继而以同样速率升温到室温28℃左右,测量分析相关数据。重复1次冷冻-复温过程记录相关数据,通过分析实验过程中的热流曲线,对比冷冻过程中的相变潜热和结冰点温度,分析不同复温速率和反复冷冻-复温过程对葡萄试样结冰过程的影响。
实验得到不同速率下冷冻过程的热流曲线, 以2、8℃/min速率下的冷冻过程热流曲线为例,如图6所示。
图6 重复1次冷冻过程的热流曲线Fig.6 Heat flow curve of repeat freezing process
从图7、8可以发现:冷冻和重复1次冷冻条件对比葡萄试样的相变潜热和结冰点温度的变化非常小,相变潜热在2、6、8、11、14、20℃/min的速率下分别变化了6.7%、3.4%、1.4%、1.5%、1.3%、4.2%。结冰点温度在2、6、8、11、14、20℃/min的速率下分别变化了0.4%、1.2%、0.8%、0.8%、1.1%、0.9%。

图7 冷冻过程与重复1次冷冻过程中结冰点温度的对比Fig.7 Freezing temperature contrast during repeated freezing process and freezing process
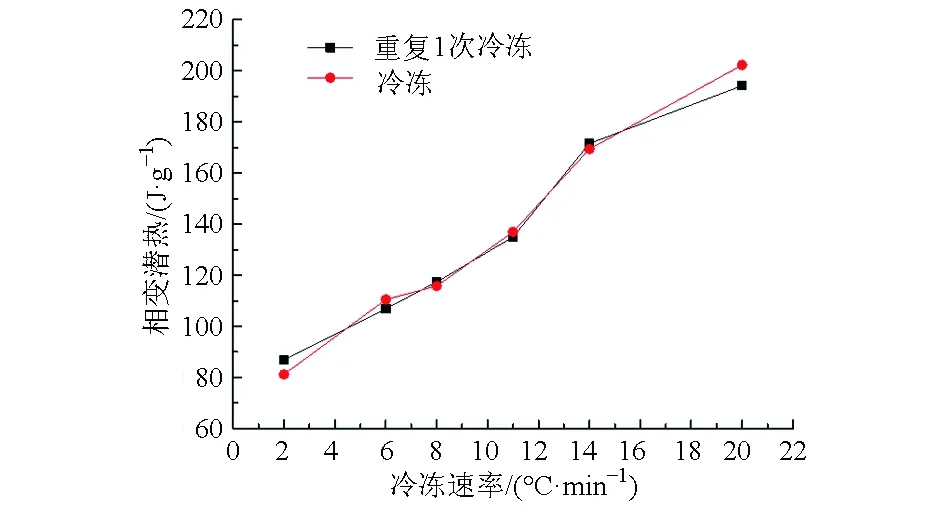
图8 冷冻过程与重复1次冷冻过程相变潜热对比Fig.8 Heat flow contrast during repeated freezing process and freezing process
2.3复温过程结果与分析
2.3.1不同复温速率对葡萄试样物性的影响
为了探究不同复温速率在冷冻-复温和重复1次冷冻-复温过程中对葡萄试样冰晶融化过程的影响,对葡萄试样先以14℃/min的速率降至-40℃,再分别以2、6、8、10、14、20℃/min的速率复温到室温28℃左右,记录相关数据。重复1次冷冻-复温过程并记录相关数据,通过分析实验过程中的热流曲线,分析不同复温速率和重复1次冷冻-复温过程对葡萄试样冰晶融化过程的影响。图9是在10℃/min速率下复温得到的热流曲线。

图9 重复复温过程的热流曲线Fig.9 Heat flow curve of repeated thawing process

图10 复温过程和重复1次复温过程的热流曲线对比Fig.10 Heat flow curve contrast during repeated thawing process and thawing process
对比不同复温速率和重复1次冷冻-复温条件下,葡萄试样相变开始点即冰晶融化点的温度。从图10可知1次复温和重复复温条件下葡萄试样的冰晶融化点温度变化很小,在2、6、8、10、14、20℃/min的速率下分别变化了0.14、0.2、0.14、0.02、0.12、0.08℃。峰点温度和结束点温度变化也都小于2℃。潜热分别变化了1.4、4.9、6.4、0.3、3.2、0.7 J/g,说明重复1次冷冻-复温过程对葡萄试样的物性变化影响较小。在重复1次冷冻-复温过程中,随着复温速率的增加,相变融化点基本不变,最大变化值为0.2℃。相变峰点温度最大增加了0.94倍,结束点温度最大增加了5.07倍,潜热最大增加了1.79倍。由此可知随着复温速率的增大,相变峰点温度、相变结束点温度和相变潜热都会增大,但是相变起始点即融化点温度基本不变。
由以上分析可知,增大复温速率能有效提高冰晶融化相变过程的潜热,这是由于在复温过程中细胞间隙的冰晶首先融化,而在冰晶融化过程中0℃的水的导热系数为0.56 W/(m·K), 冰的导热系数为2.22 W/(m·K),约为水的4倍[11-14],因此首先融化的游离态的水不能有效地将热量传递到内部区域。在慢速复温的情况下葡萄试样外侧解冻层的细胞组织已经融化或由于过热度太大而使细胞膜的渗透性被破坏,而内部结冰区仍未融化,而在快速复温时葡萄试样内外层几乎同步解冻,加快了相变过程,各个部位的细胞组织温度变化很小,加快了热量的传递,因此吸收的潜热较多,这与张素文[5]在玻璃态下冻结-冻藏及其后续解冻对西兰花品质的影响研究结论一致。另外,快速复温时,细胞相变时间短,增大了冰晶融化速率,使葡萄细胞组织快速恢复持水能力,而相变过程的峰点代表最大的转换速率点,因此随着复温速率的增加,相变峰点的温度也增大。
2.3.2通过最大冰晶生成带的速率对复温后葡萄试样物性的影响

图11 2℃/min复温过程的热流曲线Fig.11 Heat flow curve of thawing process at 2℃/min
复温过程中,为了探究试样以不同速率通过最大冰晶生成带(-6~0℃)时对试样物性的影响,对葡萄试样先以14℃/min的降温速率降至-40℃,再分别以2、6、8、10、14、20℃/min的速率复温到-6℃,然后对不同速率复温后的葡萄试样再分别以1℃/min和20℃/min升温通过最大冰晶生成带(-6~0℃)。图11是在2℃/min复温速率下得到的热流曲线。

图12 1℃/min和20℃/min通过最大冰晶生成带的热流曲线对比Fig.12 Heat flow curves contrast during the largest ice crystals generated at 1℃/min and 20℃/min
图12表示以1℃/min 和20℃/min通过最大冰晶生成带-6~0℃时融化点温度、峰点温度、相变结束点温度和潜热,在不同复温速率条件下的变化过程。可以发现1次冷冻-复温和重复冷冻复温条件下葡萄试样的冰晶融化点温度变化很小,在2、6、8、10、14、20℃/min的速率下的变化都小于0.3℃。相变峰点温度变化值小于2℃,从图中可知20℃/min的相变结束点温度高于1℃/min的相变结束点温度,随着复温速率的增加分别增加了3.44、2.86、2.59、2.28、1.76、1.33℃,潜热分别增加了19.84、17.57、9.78、6.58、11.92、15.26 J/g。从中可以看出以20℃/min的速率通过最大冰晶生成带要比1℃/min速率通过最大冰晶生成带的相变结束点温度增大98%,而潜热最大增加49%。因此在最大冰晶生成带区域,提高复温速率能有效提高相变潜热值。
大部分食品在冷冻过程中,在最大冰晶生成带范围内约80%的水分形成冰晶,同时在复温过程中最大冰晶生成带内生化反应、酶反应和细菌污染都易于发生,因此在冷冻-复温过程中都应快速通过最大冰晶生成带[15-17]。以20℃/min的速率通过最大冰晶生成带要比1℃/min速率通过最大冰晶生成带的相变结束点温度增大了98%,而潜热最大增加了49%。
2.3.3不同速率梯度对复温后葡萄试样物性的影响
复温过程中,为了探究试样以阶梯式速率复温对葡萄试样复温后物性的影响,对葡萄试样先以20℃/min的降温速率降至-40℃,再分别以2、6、8、10、14、20℃/min的速率复温到-15℃,然后对不同速率复温后的葡萄试样分别阶梯增加速率和阶梯降低速率,如在-40~-15℃区间以2℃/min复温,对应的阶梯增加速率为以4℃/min复温到28℃,对应的阶梯降低速率为以1.5℃/min复温到28℃,具体过程如表2所示。
图13表示的是在2℃/min速率下由-40℃复温到-15℃,再分别以增加速率4℃/min、降低速率1.5℃/min,由-15℃复温到28℃得到的热流曲线图。
对比增加速率梯度和降低速率梯度的不同复温速率在-15~28℃的相变过程变化,可以发现在增加速率梯度和降低速率梯度的条件下相变过程的融化点温度变化和相变结束点温度变化都小于1℃。从图14中可知增加速率梯度的相变峰点温度高于降低速率梯度的相变峰点温度,在2、6、8、10、14、20℃/min的速率下分别变化了3.06、3.33、3.69、2.65、1.39、3.05℃。可知增加速率梯度能有效提高相变峰点温度。相变潜热分别增加了43.0、43.1、45.9、52.7、54.3、45.1 J/g。可知增加速率梯度能有效提高相变潜热,增加速率梯度比降低速率梯度的潜热最大增加了39.4%,相变峰点温度最大增加了26.3%。因此增加速率梯度能有效提高相变潜热。

表2 不同速率梯度下的复温过程Tab.2 Different rate gradients in process of thawing

图13 不同速率梯度复温过程的热流曲线Fig.13 Heat flow curves of different rates of gradient during thawing process

图14 不同速率梯度复温过程的热流曲线对比Fig.14 Heat flow curve contrast during the thawing process with different rate gradients
3 结论
(1)在冷冻过程中相变潜热随降温速率的增大而增大,冰点温度随降温速率的增大而降低,在降温速率由2℃/min到20℃/min变化过程中,相变潜热增大了1.23倍,冰点降低了38.4%。
(2)随着复温速率增加,相变峰点温度、相变结束点温度和相变潜热都会增大,复温速率由2℃/min变化到20℃/min的过程中相变潜热变化了1.79倍,相变峰点温度变化了0.94倍,而相变结束点温度变化了5.07倍,但是相变起始点即融化点温度基本不变。
(3)重复1次冷冻-复温实验对葡萄试样的相变潜热、相变峰点温度以及相变结束点温度的影响都较小,变化率都小于0.75倍。
1THIRUMALA S, HUANG Changjiang, DONG Qiaoxiang, et al. A theoretically estimated optimal cooling rate for the cryopreservation of sperm cells fromalive-bearing fish, the green swordtailXiphophorushelleri[J].Theriogenology,2005, 63(9):2395-2415.
2MORI Shoji, CHOI Jeunghwan, DEVIEDDY R V, et al. Calorimetric measurement of water transport and intra-cellular ice formation during freezing in cell suspensions[J]. Cryobiology, 2012, 65(3):242-255.
3DEVIREDDY R V, RAH A D, BISCHOF J C. Measurement of water transport during freezing in cell suspensions using a differential scanning calorimeter[J]. Cryobiology, 1998, 36(2): 124-155.
4SORGENTINI D A. Comparative study of structural characteristics and thermal behavior of whey and isolate soybean proteins[J]. Journal of Food Biochemistry,1999,23(5):489-507.
5张素文. 玻璃态下冻结_冻藏及其后续解冻对西兰花品质的影响研究[D].无锡:江南大学,2007.
ZHANG Suwen. The study of the influence of freezing, storage and thaw on the quality of glassy broccoli[D].Wuxi:Jiangnan University,2007. (in Chinese)
6樊振江,高愿军,常广双,等.冻结温度对猕猴桃果丁细胞结构和质构特性的影响[J].农产品加工,2008(3):52-54.
FAN Zhenjiang, GAO Yuanjun, CHANG Guangshuang,et al. Effects of different freezing temperatures on the cells structures and texture of kiwi fruit dices [J]. Academic Periodical of Farm Products Processing,2008(3):52-54. (in Chinese)
7张哲,赵静,田津津,等. 冷冻-复温过程中葡萄细胞结晶变化的研究 [J]. 农业机械学报, 2016,47(5):211-217.
ZHANG Zhe, ZHAO Jing, TIAN Jinjin,et al. Research on crystallization change of grape cells during freezing-thawing process[J]. Transactions of the Chinese Society for Agricultural Machinery, 2016,47(5):211-217. (in Chinese)
8曾彦彰. 预处理生物材料的差示扫描量热学研究[D]. 北京:中国科学院研究生院,2009.
ZENG Yanzhang. Differential scanning calorimetric study on pretreated biological materials[D].Beijing:Chinese Academy of Sciences,2009. (in Chinese)
9苏芸. Al2O3-H2O纳米流体热物性及相变蓄冷特性的研究[D]. 天津:天津商业大学, 2013.
SU Yun. Study on thermophysical and phase change cold-storage characteristics of Al2O3-H2O nano fluids [D]. Tianjin: Tianjin University of Commerce, 2013. (in Chinese)
10易小红. 火龙果真空冷冻干燥传热传质理论与实验研究[D].天津:天津商业大学, 2008.
YI Xiaohong. Theoretical and experimental study on heat and mass transfer of firedrake fruit’s vacuum freeze-drying [D]. Tianjin: Tianjin University of Commerce,2008. (in Chinese)
11潘见.农产品流变热力学特性初探草莓组织的冰点解冻点和热分解点测定[J].江苏工学院学报,1987,8(4):18-24.
PAN Jian. Approach to rheo thermodynamic characteristics of farm products [J].Journal of Jiangsu Institute of Technology,1987,8(4):18-24. (in Chinese)
12ACKERJ P, CROTEAU I. Pre- and post-thaw assessment of intracellular ice formation[J]. Journal of Microscopy, 2004, 215(2):131-138.
13SHIURBA R. Freeze-substitution origins and applications [J]. International Review of Cytology, 2001, 206:42-47.
14ACKER J P, MCGANN L E. Cell-cell contact affects membrane integrity after intracellular freezing[J]. Cryobiology, 2000, 40(1):54-63.
15MYERS S P, PITT R E, LYNCH D V, et al. Characterization of intracellular ice formation in drosophila melanogaster embryos [J]. Cryobiology, 1989,26(5):472-484.
16KORBER C, ENGLICH S, RAU G. Intracellular ice formation cryomicroscopical observation and calorimetric measurement [J]. Journal of Microscopy, 1991,161(2):313-325.
17KLEINHANS F W, GUENTHER J F, ROBERTS D M, et al. Analysis of intracellular ice nucleation inXenopusoocytes by deferential scanning calorimetry [J].Cryobiology, 2005, 52(1):128-138.
Research on Phase Transition of Grape during Freezing-Thawing Process
Zhang Zhe1Hao Junjie1Zhao Jing1Wang Huaiwen1Ji Hongwei1Zhang Ping2
(1.TianjinKeyLaboratoryofRefrigerationTechnology,TianjinUniversityofCommerce,Tianjin300134,China2.TianjinKeyLaboratoryofPostharvestPhysiologyandStorageofAgriculturalProducts,NationalEngineeringandTechnologyResearchCenterforPreservationofAgriculturalProducts,Tianjin300384,China)
With the improvement of people’s living standard, people are more and more concerned about the quality of fruits and vegetables during freezing and cold storage. However, fruits and vegetables are constant in the process of freezing-thawing during freezing and cold storage. Therefore, it is very important to research the phase transition of fruits and vegetables during the freezing-thawing process. The phase transition of fruits and vegetables in the freezing-thawing process was studied by using red grape as the test object. Grape pulp was tested with different rates, frequencies, time intervals, rate gradients and the rates through maximum ice crystal generation zone during the freezing-thawing process by DSC. By analyzing the heat flow curve in the experiment, the effect of freezing-thawing process on grape sample under different conditions was found. The greater the freezing rate was, the lower the freezing point temperature was, the shorter the time during the freezing process was. With the increase of the rate, the peak temperature of phase transition, the end point of phase transition temperature and the latent heat of phase change all increased during the process of thawing. In the process of temperature change from 2℃/min to 20℃/min, the latent heat of phase change increased 1.79 times, the peak temperature of phase transition increased 0.94 times, and the end point of phase transition temperature increased 5.07 times. The study provides a basis of quality recovery after freezing-thawing process for fruits and vegetables.
grape pulp; freezing-thawing; phase transition; DSC system
10.6041/j.issn.1000-1298.2016.09.034
2016-06-15
2016-07-07
国家自然科学基金项目(11572223)和天津市自然科学基金重点项目(14JCZDJC34600、 15JCZDJC34200)
张哲(1975—),男,副教授,主要从事食品冷链研究,E-mail: zhangzhe@tjcu.edu.cn
TS201.1; TQ026
A
1000-1298(2016)09-0241-08
