珍稀克隆植物北极花有性繁殖及其对资源的分配和利用
2016-07-23翟雅芯王玥魏仙仙何爽张爱勤
翟雅芯,王玥,魏仙仙,何爽,张爱勤
(新疆大学生命科学与技术学院,乌鲁木齐 830046)
珍稀克隆植物北极花有性繁殖及其对资源的分配和利用
翟雅芯,王玥,魏仙仙,何爽,张爱勤
(新疆大学生命科学与技术学院,乌鲁木齐830046)
摘要:【目的】有性繁殖使克隆植物具有潜在适应环境变化的能力,对植物生活史具有重要影响。【方法】为了深入理解植物有性繁殖各构成因素之间的相互关系及对资源的分配和利用情况,通过野外观测及室内测定,研究珍稀克隆植物北极花有性繁殖与相关构成因素。【结果】北极花分株长达(60.14 ± 5.70)cm,各分株的构件参数营养枝数量、有性枝数量和分株果实数分别为(8.23 ± 1.18)枝、(2.50 ± 0.30)枝和(1.47 ± 0.29)个。有性繁殖所占分株生物量比例为7.06% ± 1.30%。在分株水平上,有性繁殖适合度的直接影响因素为有性分枝数量,不受分株长度和生物量的影响;而营养枝作为克隆株的同化器官,可间接的影响有性繁殖。在居群水平上,斑块内果实数与有性分枝数量、结实率的相关系数分别为0.849、0.847,均呈极显著相关(P<0.01);而与斑块大小(面积)无相关性。【结论】北极花具有相对低的有性繁殖状态,有性繁殖的衡量参数果实数仅与有性分枝数量和结实率相关,而表现出与对繁殖适合度有较大贡献的其他性状不相关。依据遗传不育进化的流行假说,如果北极花的有性繁殖对维持居群遗传多样性的贡献率过低,那么其有性繁殖有通过不育突变而衰退的风险。
关键词:北极花(Linnaea borealis L.);克隆植物;有性繁殖;构件参数;相关分析
0引 言
【研究意义】大多数克隆植物兼具有性和无性两种繁殖方式,并对居群的遗传结构产生重要影响。有性繁殖可通过配子体的有性融合、染色体分离和等位基因重组等产生遗传变异[1],以增加植物潜在的适应环境变化的能力。但不同克隆植物之间有性繁殖和实生苗的更新能力差异很大,一些植物因克隆繁殖甚至放弃了有性过程,进而导致物种遗传多样性的降低和对环境适应力的下降[2-3]。随着珍稀物种孤立小种群因有性繁殖衰退而导致濒危的报道不断增多[4-6],植物有性繁殖及其相关因素的研究成为许多克隆植物生活史研究的重要内容。植物的有性繁殖受多种因素的影响:如生境破碎化使物种以小居群的形式存在,居群间基因流中断,近交比例过高,两种繁殖方式间的资源权衡,及由此产生花粉输出下降的雄性适合度代价和干扰结实产生的雌性适合度代价。尤其像高山、极地等一些特殊环境,传粉者数量有限、结实率过低、或者结实后种子难以萌发等问题也成为植物有性繁殖的重要制约因素[4,7-10]。【前人研究进展】北极花(LinnaeaborealisL.)也叫林奈木,以分布在北半球高纬度地区而命名,也因优美的花型和作为植物界最小的灌木,被冠以著名分类学家的名字林奈,而深受喜爱。作为高纬度寒冷地区的一个常绿、匍匐小灌木,具克隆生长习性的一个珍稀物种,由于居群的片断化导致有性繁殖受阻[12-15]。在苏格兰等地已被定为濒危物种[16],我国目前将其作为珍稀物种记载[17]。目前针对克隆植物有性繁殖的研究主要集中在植物两种繁殖方式的进化权衡、克隆生长在花粉传递与交配系统方面对有性繁殖所产生的影响[11]。【本研究切入点】自然状态下,有性繁殖与其构成因素的相互作用及各因素对资源的利用和分配情况还缺乏研究。严酷的生境集有性、无性繁殖于一体,使北极花成为进行植物繁育系统多样性研究和评估濒危克隆植物有性繁殖状态,分析濒危机制的较好材料。【拟解决的关键问题】研究北极花的有性繁殖及相关构成因素如分株果实数、分株长度、有性分枝数、无性分枝数、分株生物量大小、结实率等,为揭示克隆植物的有性繁殖状态和多样的繁育系统类型积累新的数据。
1材料与方法
1.1材 料
观测点位于86°55′E~87°03′E,48°36′N~48°42′N的喀纳斯自然保护区海拔1 500~2 000 m的针叶林下。北极花为游击型克隆构型,以每年产生6~8对对生叶片、长度30~50 cm的方式向前延伸,具较强的克隆生长力。有性和无性两类垂直短枝着生在对生叶片的叶腋处。花序生于有性枝的顶端,具2花,双悬垂,花果期6~8月。图1
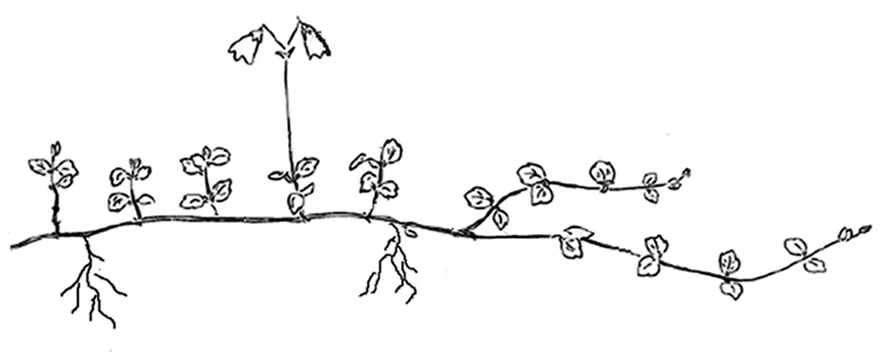
图1 北极花游击型克隆分株
1.2方 法
1.2.1分株水平上有性繁殖与构成因素的测定及各构成因素的相关分析
在北极花结实末期,以间隔5 m左右的距离为准,在30个不同的克隆斑块内随机取匍匐茎分株46个,对所有分株的长度分别进行测量。同时,随机对30个分株进行有性枝、无性枝数量及分株果实数的统计。并对分株果实数与各构成因素进行相关分析。
1.2.2分株生物量及有性繁殖分配比例的测定及相关分析
在北极花结实末期,间隔一定距离在不同的克隆斑块内随机选取30个匍匐茎分株,清理干净泥土后,将各分株分为有性繁殖构件(所有有性分枝及其上边的果实)、无性分枝、及分株匍匐茎三部分,在80℃恒温箱中烘72 h、分别称干重,后进行各分株生物量和有性繁殖分配比例的统计。
1.2.3居群水平上有性繁殖格局及其与构成因素的相关分析
在结实期随机选定不同面积大小的样方(克隆斑块)30个,对其面积进行测定;统计单位面积内有性分枝数量和果实数,计算结实率;并进行斑块内果实数与面积大小等各构成因素的相关性分析。
1.2.4种子与幼苗的补充及更新
为了从后代适合度方面衡量有性繁殖的贡献率,随机选定不同面积大小的克隆斑块10个,进行如下观测:(1)对斑块内实生苗数量进行统计;(2)对居群中当年新形成的果实及种子进行发育状况的跟踪观察。(3)对斑块内不同时期的结实率进行统计。结实率为克隆斑块内所有果实数与单花数的比。
1.3数据处理
用SPSS 17.0,Excel 2007软件进行数据处理、分析和绘制图表。首先用One-Sample Kolmogorov-Smirnov Test进行正态性检验,经检验各组数据均符合正态分布后,再进行相关数据分析。所有统计数据均用平均值±标准误表示。
2结果与分析
2.1分株水平上有性繁殖与构成因素
研究表明,北极花克隆分株的长度变化幅度较大,最长的达145.60 cm,最短的仅8.55 cm;平均长度为(60.14 ± 5.70 )cm,为典型的游击型克隆构型。分株上有性和无性枝条从叶腋处伸出,随机分布在匍匐茎上,平均每分株有营养枝(8.23 ± 1.18)枝(最大值为22枝,最小值为0);有性枝(2.50 ± 0.30)枝(最大值为8枝,最小值为0)。每分株上有效果实数最大值为6个,最小值为0,平均为(1.47 ± 0.29)个。
从各相关参数的测定值看,北极花分株较长,但各分株的构件参数如营养枝数量、有性枝数量、分株上果实数等较低,且各参数变异幅度较大。尤其是有性繁殖的构成因素如有性枝数量、果实数及生物量分配比例等显得尤为低下。表1
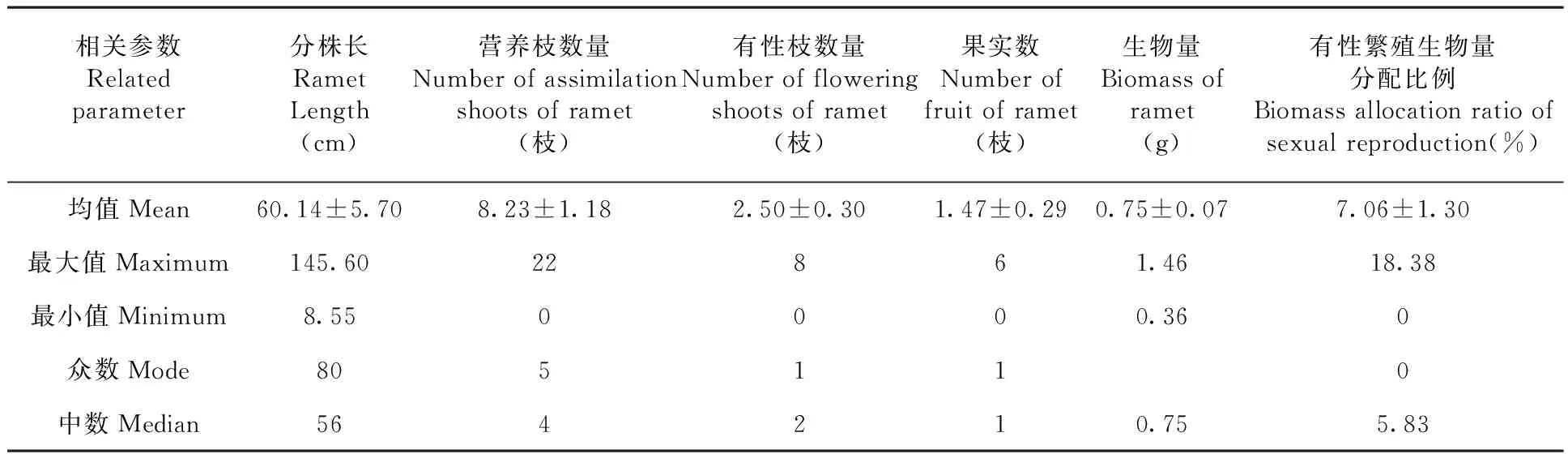
表1 分株水平上北极花果实数量与构成因素的测定值(平均值±标准误)
2.2分株生物量及有性繁殖分配比例
研究表明,北极花分株生物量为(0.75 ± 0.07)g,分株上营养枝和分株匍匐茎所占的生物量比例分别为45.32% ± 3.56%,47.62% ± 3.33%;远高于有性繁殖7.06% ± 1.30%的生物量分配比例。
北极花被称为世界上最小的小灌木,从分株生物量的大小就可看出其细弱的株型。但较长的分株匍匐茎,说明了匍匐茎及其附着的根系在克隆株营养物质储藏、资源的获取、基株生长寿命的维持方面所具的重要功能。在分株各构件参数中,分株匍匐茎生物量最高也说明了这一现象。图2
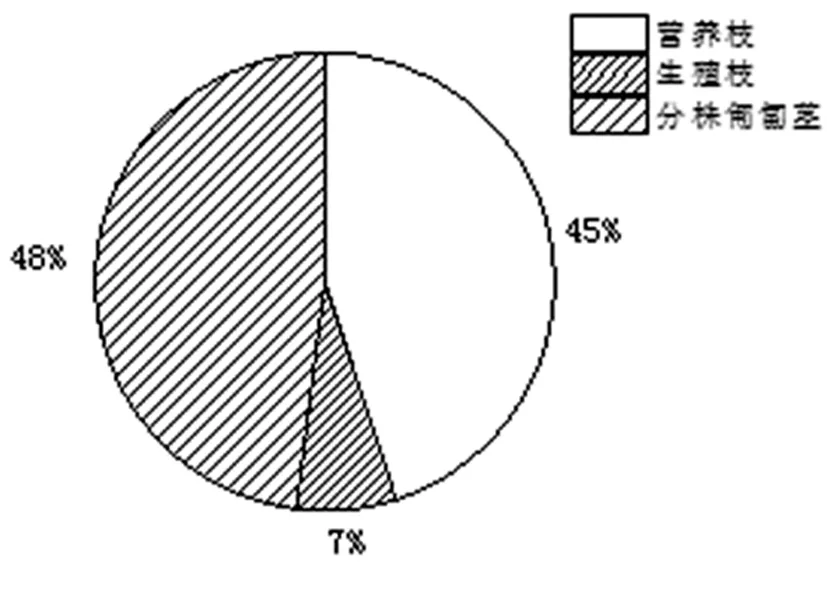
图2 北极花分株各构件生物量分配比例
2.3分株水平上北极花有性繁殖与构成因素的相关性
研究表明,不同分株长度的果实数与有性枝数量的相关系数是0.389,呈显著相关(P<0.05);与分株长度、分株上无性枝数量、分株的生物量无显著相关性。而在各构成因素中,有性枝数量与营养枝数量的相关系数是0.422,为显著相关(P<0.05);分株长度与营养枝数量的相关系数为0.821,呈极显著正相关(P<0.01);与有性枝数量、生物量和果实数相关系数分别为0.291、0.032和0.017,相关性均不显著。
从各构成因素的相关性看,有性繁殖适合度的直接影响因素为有性分枝数量,而营养枝为克隆株的同化器官,是分株光合产物的主要提供者,与有性枝的数量呈正相关,可间接的影响有性繁殖。表2
2.4克隆斑块内有性繁殖格局及果实数与构成因素的相关性
研究表明,北极花斑块内的果实数与有性分枝数量、结实率均呈极显著正相关(P<0.01),其相关系数分别为0.849、0.847;而与斑块面积的相关性不显著,表明在斑块内,北极花的果实数、有性分枝数呈明显的非均匀分布,有性繁殖适合度主要受斑块内有性枝数量和结实率影响。表3
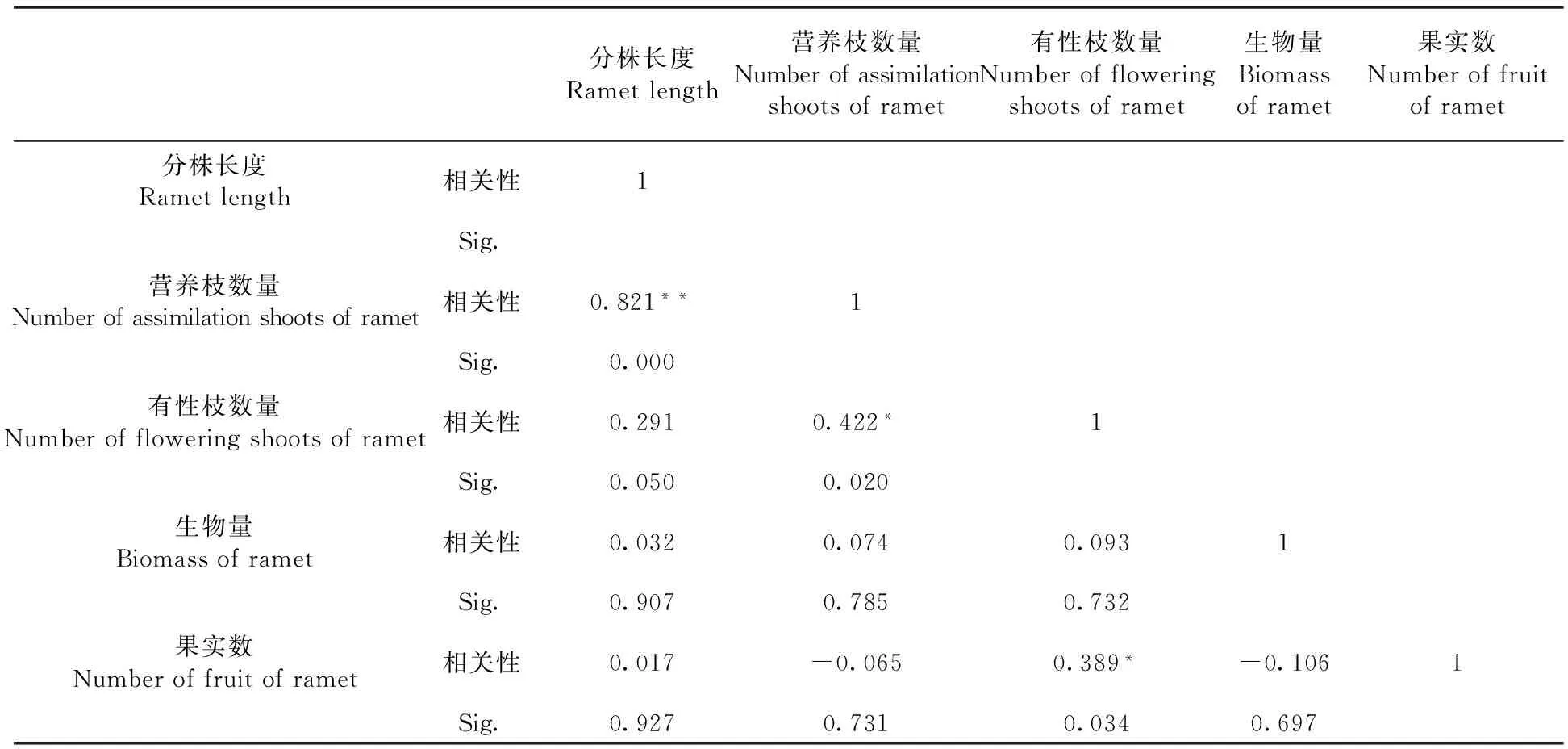
表2 分株水平上北极花果实数与构成因素的相关性
注:**在0.01水平(双侧)上显著相关。*在0.05水平(双侧)上显著相关
Note:**the 0.01 level. (bilateral). A significant correlation at 0.05 level (bilateral) significant correlation
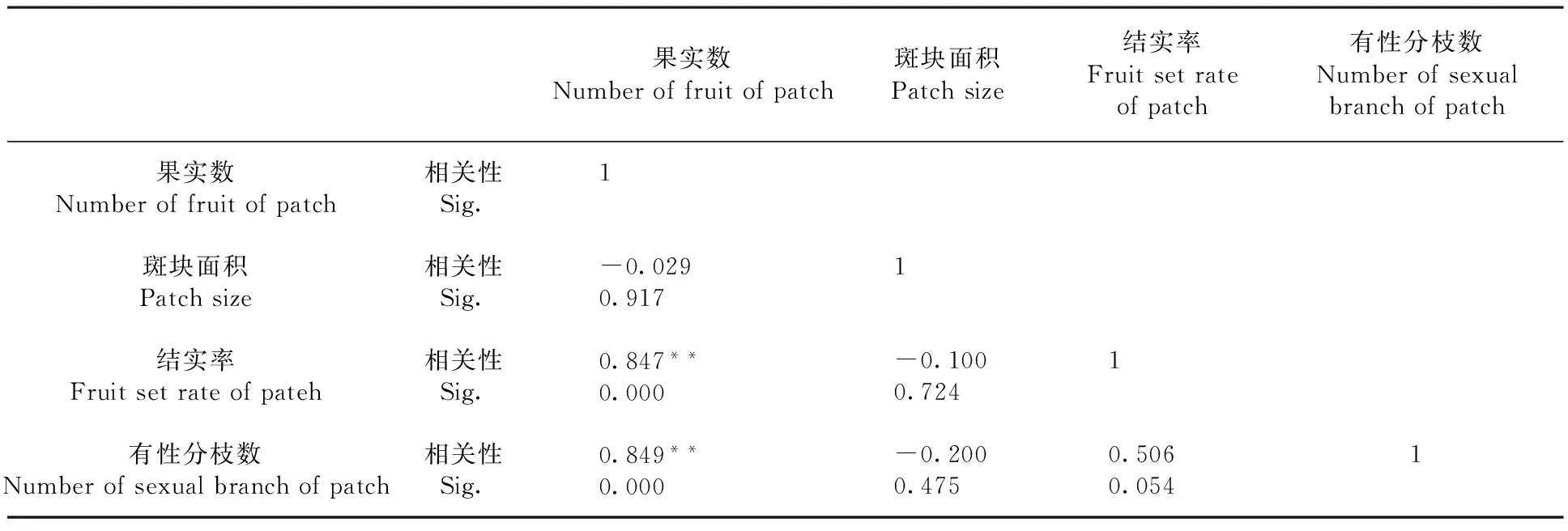
表3 北极花克隆斑块内果实数与斑块面积大小、结实率和有性枝数量的相关性
注:**在0.01水平(双侧)上显著相关
Note:**at the 0.01 level (bilateral),a significant correlation
2.5种子与幼苗的补充及更新
自然状态下北极花的胚珠有10多枚,但只有1枚能够发育,授粉后10 d的胚珠在大小上仅略微膨大,幼果大小变化也不明显。结实初期(自然授粉后半个月)结实率高达82.18% ± 1.48%,但后期逐步脱落。自然授粉后1个月观测,果实及种子仍为绿色,结实率为35.64% ± 2.20%。当年果实完全脱落,无宿存现象。对10个大小不等的克隆斑块进行幼苗补充状况的调查,不见幼苗的更新。在居群的繁殖与更新中有性繁殖的后代适合度较低。
3讨 论
3.1北极花的繁殖模式
克隆繁殖使得个体在有效摄取资源、规避风险方面具有重要作用,但是在资源有限的情况下,旺盛的克隆生长极易降低有性繁殖的资源投入。通常“游击型”克隆植物能有效利用异质生境中的资源,具有较强的“觅食行为”,但有性繁殖能力较低[18-20]。北极花居群的繁殖模式支持了这一观点。
北极花的分株长度达(60.14 ± 5.70)cm,但其分枝数量并不多,且在单株水平上有性枝数量(2.50 ± 0.30)枝明显低于营养枝数量(8.23 ± 1.18)枝,在分株生物量统计中,有性繁殖的生物量分配比例仅占7.06% ± 1.30%,远低于分株匍匐茎和营养枝的生物量,后者的生物量之和占90%以上。自然界中,植物的生殖分配比例是相对稳定的,通常一年生植物生殖分配比例达20%~40%,多年生植物占20%左右,人工选择下的栽培植物可达40%左右[21]。相比之下,北极花的有性繁殖分配比例要低得多。
居群中实生苗的补充状况是衡量有性繁殖适合度的主要参数之一,尽管统计显示每分株有果实(1.47 ± 0.29)个,但在野外观测中几乎没有看到实生苗的补充。从两种繁殖方式的适合度看,北极花居群的扩展更多的依赖于分株匍匐茎的伸长和营养枝的分化。
3.2北极花的有性繁殖
植物繁殖与最适生活史格局的形成通常受植物资源的分配模式,尤其是繁殖过程中资源的分配模式影响[22]。生活史理论预测:不同生活史类型的植物各性状之间的资源分配存在着“此消彼长”的权衡关系[11]。但这种权衡又受资源条件、生长时期和遗传因素等的影响而表现各异。北极花在分株和居群水平上,其有性繁殖的衡量参数果实数仅与有性分枝数量和结实率相关,而表现出与对繁殖适合度有较大贡献的其他性状不相关,没表现出明显的权衡关系。权衡关系的出现取决于可利用的资源的量,只有当资源存在限制,而且依赖于相同的限制资源时,才能表现出来。
北极花过低的有性繁殖力可能在资源分配方面不足以对分株产生影响,或者与其他性状之间依赖于不同的限制性资源,进而表现出与其他性状的不相关。
有性繁殖对克隆植物居群遗传结构贡献的研究在不同类群中差异很大。一些研究表明较低的有性繁殖使得居群处于濒危状态[4-6];而又有一些研究则认为能够产生大量种子并不意味着是以有性繁殖为主,如果幼苗成功更新的条件存在,不频繁的种子生产也可能维持高的遗传多样性[23,24]。为此,多种机制被提出用于解释有性繁殖对克隆植物遗传结构的贡献,其中种群幼苗补充机制假说认为:克隆植物居群遗传多样性的维持可通过建群早期的幼苗补充、发育过程中的重复幼苗补充和通过干扰发生的阵发式幼苗补充三种途径来实现。其中,初期幼苗补充和阵发式幼苗补充在居群中都不会看到有高的幼苗补充[25,26]。这一假说较好的解释了一些居群有着高的遗传多样性,却没有高的幼苗补充的现象。北极花属于哪种情况还有待于对其进一步研究。
观测点位于北极花亚欧地理分布区的边缘,依据遗传不育进化的流行假说:居群中一些对适合度不再有贡献的性状,往往可能会通过突变而逐步退化[27]。北极花居群过低的有性繁殖率如果对居群适合度不再有贡献或贡献率过低,那么其有性繁殖就有通过不育突变累积而退化的风险。
4结 论
4.1北极花具有较强的克隆生长力和相对低的有性繁殖状态,居群的扩展更多依赖于分株匍匐茎的伸长和营养枝的分化。
4.2有性繁殖的衡量参数果实数仅与有性分枝数量和结实率相关,过低的有性繁殖力可能在资源分配方面不足以对分株产生影响,或者与其他性状之间依赖于不同的限制性资源,进而表现出与对繁殖适合度有较大贡献的其他性状不相关。
4.3依据遗传不育进化的流行假说,如果北极花的有性繁殖对维持居群遗传多样性的贡献率过低,那么其有性繁殖有通过不育突变而衰退的风险。这也是很多物种濒危的重要原因之一。
参考文献(References)
[1]张大勇. 植物生活史进化与繁殖生态学[M]. 北京: 科学出版社, 2004.
ZHANG Da-yong. (2004).Lifehistoryevolutionandreproductionecologyinplants[M]. Beijing: Science Press. (in Chinese)
[2] Eriksson, O. (1997).Clonallifehistoriesandtheevolutionofseedrecruitment. [C]//. In: de Kroosn H & van Groenendael (eds). The ecology and evolution of clonal plants. Leiden: Backhuys Publishers. pp. 66.
[3] 张玉芬, 张大勇. 克隆植物的有性与无性繁殖对策[J]. 植物生态学报, 2006, 30(1): 174 - 183.
ZHANG Yu -fen, ZHANG Da-yong. (2006). Asexual and sexual reproductive strategies in clonal plants [J].JournalofPlantEcology, 30(1): 174 - 183. (in Chinese)
[4] Fischer, M., Hock, M., & Paschke, M. (2003). Low genetic variation reduces cross-compatibility and offspring fitness in populations of a narrow endemic plant with a self-incompatibility system.ConservationGenetics, 4(3):325-336.
[5] Aigner, P. A. (2004). Ecological and genetic effects on demographic processes: pollination, clonality and seed production in dithyrea maritima.BiologicalConservation, 116 (1):27-34.
[6] Scobie, A. R., & Wilcock, C. C. (2009). Limited mate availability decreases reproductive success of fragmented populations of linnaea borealis, a rare, clonal self-incompatible plant.AnnalsofBotany, 103 (6):835-846.
[7] Andrew, G. (1999). Paternal bottlenecks in fragmented populations of the grassland daisy rutidosis leptorrhynchoides.GeneticsResearch, 73:111-117.
[8] Willi, Y., Van, B. J., & Fischer, M. (2005). A threefold genetic allee effect: population size affects cross-compatibility, inbreeding depression and drift load in the self-incompatible ranunculus reptans.Genetics,169 (4):2,255-2,265.
[9] Willi, Y., Buskirk, J. V., & Hoffmann, A. A. (2006). Limits to the adaptive potential of small populations.AnnualReviewofEcologyEvolution&Systematics, 37:433-458.
[10] Totlsand. (2001). Environment-dependent pollen limitation and selection on floral traits in an alpine species.Ecology, (82): 2,233-2,244.
[11] 董鸣. 克隆植物生态学[M]. 科学出版社, 2011:67-68.
DONG Ming. (2011).ClonalPlantEcology[M]. Beijing: Science Press:67-68. (in Chinese)
[12] Wilcock, C. C., & Jennings, S. B. (1999). Partner limitation and restoration of sexual reproduction in the clonal dwarf shrub linnaea borealis, l. (caprifoliaceae).UniversityCollegeLondon, 208 (1):76-86.
[13] Scobie, A. R., & Wilcock, C. C. (2009). Limited mate availability decreases reproductive success of fragmented populations of linnaea borealis, a rare, clonal self-incompatible plant.AnnalsofBotany, 103 (6):835-846.
[14] Zhang, A. Q., Xiong, Y. Z., Huang, S. Q. (2014). Maintenance of self-incompatibility in peripheral populations of a circumboreal woodland subshrub.AoBPlants, 6(3):141-145
[15]侯真珍, 何爽, 王风雷, 等. 珍稀克隆植物北极花有性繁殖动态及影响因素[J]. 生态学杂志,2013,(12):3 167-3 172.
HOU Zhen-zhen, HE Shuang, WANG Feng-lei, et al. (2013). Dynamics of rare clonal plant Linnaea borealis L. sexual reproduction and related affecting factors [J].ChineseJournalofEcology, (12):3,167-3,172. (in Chinese)
[16] Neiland, M. R. M., Wilcock, C. C. (1997). Reproductive ecology and conservation of rare plant species in Scotland [J].BotanySocial.ScotlandNews, 68:18-19.
[17] 尹林克. 新疆珍稀濒危植物[M]. 乌鲁木齐: 新疆科学技术出版社, 2006.
YIN Lin-ke. (2006).RareendangeredendemicplantsinXinjiangofChina[M]. Urumqi: The Publish House of Xinjiang Science Technology. (in Chinese)
[18] Sculthorpe, C. D. (1967). The biology of aquatic vascular plants [M].EdwardArnold, London.
[19] Philbrick, C. T., & Les, D. H. (1996). Evolution of aquatic angiosperm reproductive systems. Bioscience, 46(11):813-826.
[20] Marcel E. Dorken +, & Eckert, C. G. (2001). Severely reduced sexual reproduction in northern populations of a clonal plant, decodon verticillatus, (lythraceae).JournalofEcology, 89 (3):339-350.
[21] 周纪伦. 植物种群生态学[M]. 北京: 高等教育出版社, 1992.
Zhou Ji-lun. (1992).PlantPopulationEcology[M]. Beijing: Higher Education Press. (in Chinese)
[22] 钟章成. 植物种群的繁殖对策[J]. 生态学杂志, 1995,14(1):37 - 42.
ZHONG Zhang-cheng. (1995). Reproductive strategies of plant populations [J].ChineseJournalofEcology, 14(1):37-42. (in Chinese)
[23] Bengtsson, B. O. (2003). Genetic variation in organisms with sexual and asexual reproduction.JournalofEvolutionaryBiology, 16(2):189-199
[24] Silvertown ,J., Franco, M., Menges, E. (1996). Interpretation of elasticity matrices as an aid to the management of plant population for conservation [J].ConservationBiology, 10:591-597.
[25] Eriksson, O. (1989). Seedling dynamics and life histories in clonal plants.Oikos, 55(2):231-238.
[26] Eriksson, O. (1993). Dynamics of genets in clonal plants.TrendsinEcologyandEvolution, 8(9):313-316.
[27] Klekowski, E. J. (1988). Progressive cross- and self-sterility associated with aging in fern clones and perhaps other plants.Heredity, 61 (2):247-253.
Fund project:NSFC- Xinjiang joint fund project (U1203102); Key Laboratory of genetic engineering of biological resources in Xinjiang (XJDX0201-2011-08); Innovative Training Program for Undergraduate of Xinjiang University(XJU-SRT-14067)
doi:10.6048/j.issn.1001-4330.2016.06.020
收稿日期(Received):2016-01-19
基金项目:NSFC-新疆联合基金项目(U1203102); 新疆生物资源基因工程重点实验室项目(XJDX0201-2011-08); 新疆大学大学生创新训练项目(XJU-SRT-14067)
作者简介:翟雅芯(1990 - ), 女, 河南郾城人, 硕士研究生, 研究方向为繁殖生态学,(E-mail)1435605732@qq.com 通讯作者(Cotresponding author):何爽(1970 - ), 女, 讲师, 研究方向为繁殖生态学, (E-mail)heshuang18@yahoo.com 张爱勤(1970 - ), 女, 河南南阳人, 教授, 硕士生导师, 研究方向为繁殖生态学, ( E-mail)zaql@sohu.com
中图分类号:Q94-1
文献标识码:A
文章编号:1001-4330(2016)06-1122-07
The Sexual Reproduction and Plant Strategies for Resource Acquisition and Mass Partitioning in Rare Clonal PlantLinnaeaborealisL.
ZHAI Ya-xin, WANG Yue, WEI Xian-xian, HE Shuang, ZHANG Ai-qin
(CollegeofLifeScienceandTechnology,XinjiangUniversity,Urumqi830046,China)
Abstract:【Objective】 Sexual reproduction has an important influence on clonal plant life history, for it increases the potential ability of clonal plant adapting to the changeable environment.【Method】To understand interactions among component factors contributing to plant sexual reproduction, and resource condition in terms of distribution and partitioning, we studied sexual reproduction effort and some related agronomical traits such as sizes of clonal ramets, no. of fruit, assimilation shoots and flowering shoots per clonal ramet, biomass allocation ratio of clonal ramets, no. of flowering shoots, fruit set ratio of different size clonal patch and so on in Linnaea borealis by observation in the wild and measurement indoor. 【Result】The results showed the clonal ramets was (60.14 ± 5.70) cm length, with assimilation shoots of (8.23 ± 1.18),flowering shoots of (2.50 ± 0.30) and fruits of (1.47 ± 0.29) per clonal ramet. The sexual reproduction fitness was affected directly by sexual branch number, and not affected by ramet length and biomass at ramet level by correlation analysis. The assimilation shoots, the assimilation organs of cloned ramets, affected indirectly sexual reproduction fitness of population by impacting on the number of flowering shoots. At population level, the number of fruit was correlated significantly with the sexual branch number and fruit set rate with the correlation coefficient of 0.849, 0.847 respectively (P<0.01), and no correlation with clonal patch size. 【Conclusion】L. borealis had relatively low sexual reproduction effort. The fruit number, the measure parameters of sexual reproduction, was only related to sexual branch number and fruit set rate, and no correlation with other traits which have greater contribution to offspring fitness such as sizes of clonal ramets, no. of assimilation shoots, ramets biomass and clonal patch size. On the basis of the evolution hypothesis of genetic sterility, the sexual reproduction would have the risk of degradation by sterility mutations in L. borealis natural populations, if the genetic diversity of population were at a lower level due to the low contribution rate of sexual reproduction.
Key words:Linnaea borealis L.; clonal plant; sexual reproduction; component parameters; correlation analysis
