钙和水杨酸对低温下黄瓜幼苗抗氧化酶同工酶的影响
2016-05-03王素平
王素平
摘要: 以黄瓜品种津绿11号为试验材料,采用聚丙烯酰胺凝胶电泳技术,研究了外源叶面喷施2 mmol/L CaCl2和1 mmol/L SA预处理对低温下黄瓜幼苗抗氧化酶同工酶的影响。结果表明:喷施2 mmol/L CaCl2对低温处理下的黄瓜幼苗SOD和POD同工酶酶带的条数和强度没有明显影响,但明显增强了CAT同工酶酶带的强度;喷施1 mmol/L SA明显增强了低温处理下黄瓜幼苗的SOD1~SOD4、POD1、POD3~POD 6、CAT1和CAT2酶带。暗示钙和SA通过影响抗氧化酶同工酶表达参与黄瓜植株低温耐性的调节。
关键词: 钙;水杨酸;低温;黄瓜;抗氧化酶;同工酶
中图分类号: S642.201 文献标志码: A 文章编号:1002-1302(2016)03-0168-03
黄瓜(Cucumis sativus L.)是我国温室栽培的主要蔬菜作物之一,黄瓜属喜温性作物,对低温反应很敏感[1],在冬春设施栽培期间经常遭受不同程度的低温胁迫,对黄瓜植株的生长、光合、碳和氮的代谢、酶活性和干物质积累产生不同程度的影响。植物体内存在的超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)等有助于植物抵御不良环境造成的活性氧伤害。Ca2+既是细胞的结构物质,又是耦联胞外信号与胞内生理生化反应的第二信使,在调控温度、水分以及盐胁迫下植物防御生理响应方面具有重要作用[2]。钙能提高逆境下植物组织或细胞的抗性,可提高辣椒(Capsicum annuum L.)幼苗的抗寒性[3],降低低温胁迫下水稻(Oryza sativa L.)幼苗的丙二醛含量[4],增强茄子(Sloanum melongena)幼苗抗膜脂过氧化能力[5]。水杨酸(salicylic acid,SA)是植物体内普遍存在的一种小分子酚类物质,在诱导植物抗病性、信号转导、激活植物过敏反应和系统获得性抗性等方面起着重要的调节作用[6]。外源SA能够缓解低温胁迫对黄瓜植株生长的抑制作用[7]。本试验以黄瓜幼苗为材料,研究了外源钙和SA预处理对低温下黄瓜幼苗抗氧化酶同工酶的影响,探讨钙和SA对黄瓜幼苗低温耐性的调节机制,以期为提高设施蔬菜作物低温耐性提供参考。
1 材料与方法
1.1 材料和处理
供试黄瓜品种为津绿11号(天津市绿丰园艺新技术开发有限公司),购自河南省商丘市种子公司。在商丘师范学院玻璃温室内培育黄瓜幼苗。黄瓜种子经温汤浸种并催芽后,于50孔穴盘中育苗,2叶1心时,定植于13 cm×12 cm软塑料营养钵中,3叶1心时将黄瓜幼苗随机分成3组,每组30株,分别进行以下不同处理。试验共设3个处理:(1)低温:将黄瓜幼苗放入人工气候箱进行低温处理,处理条件为:昼/夜温度为15 ℃/8 ℃;光/暗时间为12 h/12 h。(2)低温+2 mmol/L CaCl2:记作2 mmol/L Ca2+,在低温处理黄瓜幼苗前叶面喷施CaCl2水溶液。(3)低温+1 mmol/L SA:记作 1 mmol/L SA,在低温处理黄瓜幼苗前叶面喷施SA水溶液。CaCl2和SA处理:采用叶面喷施法进行预处理,分别于低温处理开始前60、48、36、24 h喷施4次,处理(1)同时喷施等体积的水,叶片正反面均喷到。所有处理均添加0.01%吐温。
低温处理4 d时,取黄瓜幼苗生长点下面第2张展开叶进行SOD、POD和CAT同工酶电泳。试验重复3次。
1.2 同工酶电泳
1.2.1 酶液提取 SOD、POD和CAT同工酶电泳采用同样的酶提取液。取黄瓜幼苗叶片0.4 g加1.5 mL磷酸钠缓冲液(0.05 mol/L,pH值7.8,内含0.1 mmol/L EDTA和1% PVP),冰浴研磨至匀浆。以4 ℃、12 000 r/min离心20 min,取上清液,即得SOD、POD和CAT酶液,用于SOD、POD和CAT同工酶电泳。
1.2. 2 同工酶电泳 SOD、POD和CAT聚丙烯酰胺同工酶电泳采用垂直板电泳槽进行,分离胶浓度为10%,浓缩胶浓度为3.75%,在浓缩胶中稳定电压为90 V,进入分离胶后稳定电压为200 V,3~4 h完成电泳。SOD同工酶电泳采用邻联(二)茴香胺法[8]进行染色;POD同工酶电泳采用醋酸联苯胺法[9]进行染色;CAT同工酶电泳在分离胶中加入总体积 0.5% 的可溶性淀粉,并采用H2O2-KI法[10]进行染色。
2 结果与分析
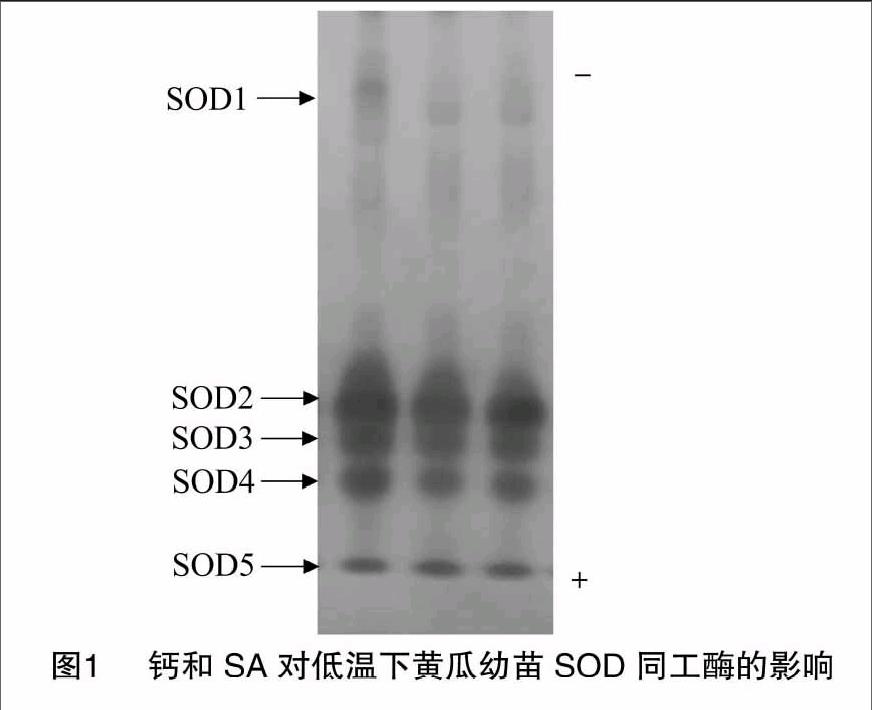
2.1 钙和SA对低温下黄瓜幼苗SOD同工酶的影响
从图1可以看出,低温处理的黄瓜幼苗SOD同工酶显现出SOD1、SOD2、SOD3、SOD4和SOD5共5条主酶带,其中SOD2、SOD4和SOD5酶带出现较早,属于强带;SOD1酶带显色较晚,着色浅,属于弱带。与低温处理相比,2 mmol/L Ca2+预处理的黄瓜幼苗SOD同工酶酶带条数不变,其着色程度与低温处理相近;1 mmol/L SA预处理明显增强了黄瓜幼苗SOD1~SOD4酶带,SOD5酶带着色程度与低温处理相近。
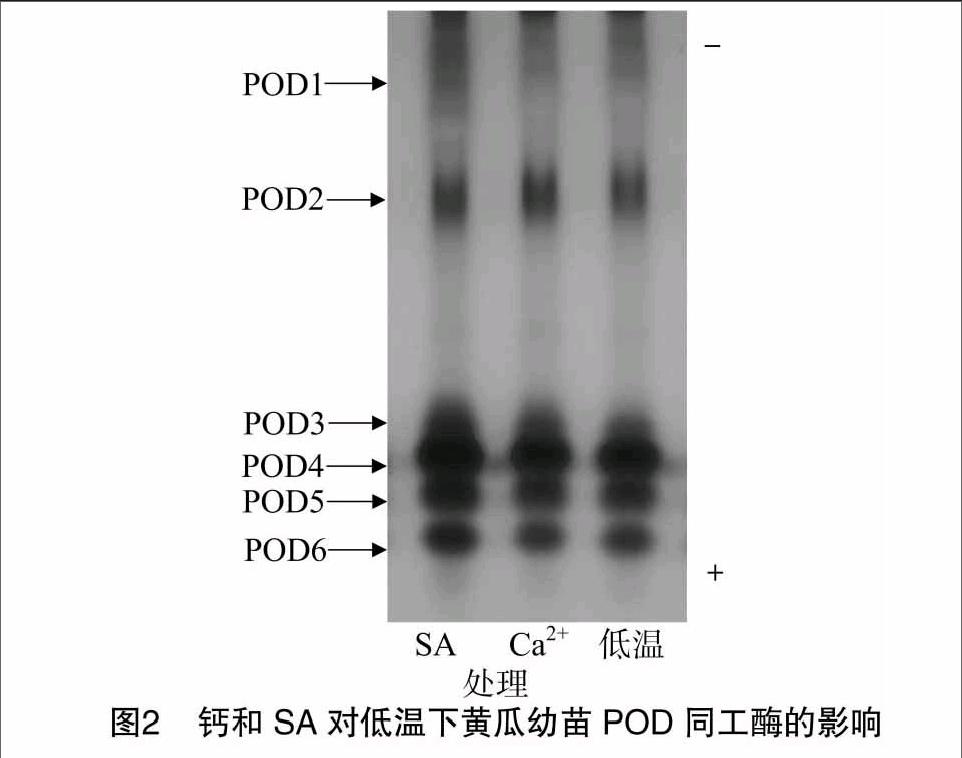
2.2 钙和SA对低温下黄瓜幼苗POD同工酶的影响
由图2可以看出,低温处理的黄瓜幼苗POD同工酶显现出6条主酶带,分别为POD1、POD2、POD3、POD4、POD5和POD6,其中POD4、POD5和POD6酶带出现较早,属于强带;POD1酶带显色较晚,着色浅,属于弱带。与低温处理相比,2 mmol/L Ca2+预处理的黄瓜幼苗POD同工酶酶带条数不变,其着色程度与低温处理相近;1 mmol/L SA预处理明显增强了黄瓜幼苗POD1、POD3~POD6酶带,POD2酶带着色程度与低温处理相近。
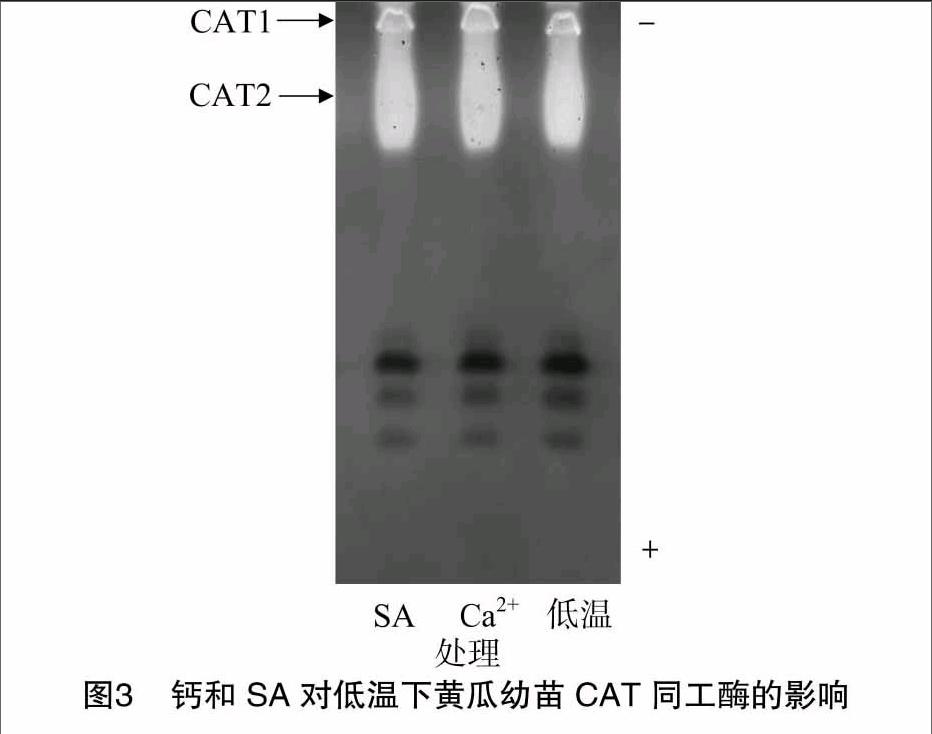
2.3 钙和SA对低温下黄瓜幼苗CAT同工酶的影响
由图3可以看出,低温处理的黄瓜幼苗CAT同工酶显现2条酶带,分别为CAT1和CAT2。与低温处理相比,2 mmol/L Ca2+和1 mmol/L SA预处理的黄瓜幼苗CAT同工酶酶带条数不变,CAT1和CAT2酶带明显增强,尤以2 mmol/L CaCl2预处理的CAT酶带为最强。
3 讨论
活性氧包括超氧阴离子(O-2 · )、H2O2、羟自由基(·OH)和单线态氧(1O2)等,植物中活性氧水平受抗氧化酶活性和抗氧化剂的调节。抗氧化酶SOD以 O-2 · 为底物进行歧化反应,起到清 O-2 · 的作用,其酶促反应产生的H2O2仍是有毒害的,H2O2可经POD和CAT等催化降解为无害的H2O。通过SOD、POD和CAT等抗氧化酶协同作用,使植株体内活性氧维持在一个较低水平,减少对细胞的伤害。同工酶的功能在生理上表现为对代谢的调节作用,同工酶分析是认识基因存在和表达的工具[11]。
Ca2+能维持细胞壁、细胞膜及膜蛋白的稳定性,在植物感应、传递抗寒信号、产生生理响应过程中起着重要作用[12-13]。Ca2+可以提高低温胁迫下黄瓜[14]、辣椒[3]、番茄(Lycopersicon esculentum Mill.)[15]、甜瓜(Cucumis melo L.)[16]和棉花(Gossypium spp.)[17]幼苗叶片SOD、POD、CAT活性,降低丙二醛含量,防止膜脂过氧化,维持膜完整性。本试验中,叶面喷施 2 mmol/L CaCl2对低温处理下的黄瓜幼苗SOD和POD同工酶酶带的条数和强度没有明显影响,但明显增强了CAT同工酶酶带的强度。表明Ca2+通过影响抗氧化酶同工酶表达参与黄瓜植株低温耐性的调节。王芳等采用CaCl2营养液浇灌玉米(Zea mays L.)幼苗[18],姜秀梅等采用CaCl2叶面喷施辣椒幼苗,其研究结果均以10 mmol/L CaCl2处理缓解低温伤害的效果最为明显[19]。李晓明等则采用0.5 mmol/L CaCl2叶面喷施低温下黄瓜幼苗[14]。由此可知,由于采用的植物材料和品种不同,Ca2+处理的方法、浓度、次数和时间不同,低温处理的强度和时间长短不同,导致研究结果存在差异。
SA被认为是植物在胁迫反应中产生的一种信号分子[20]。SA可能直接作为一种抗氧化剂淬灭活性氧或者通过激活抗氧化剂的活性调节氧化还原平衡[21-22],通过增加维生素C和谷胱甘肽的含量,降低锰胁迫对黄瓜植株造成的氧化伤害[23],通过调节抗氧化代谢促进盐胁迫下蔬菜作物的生长[24]。马德华等对黄瓜幼苗进行低温锻炼,可使叶片游离态SA含量增加2.5倍以上,表明SA可能与植物的抗低温胁迫有关[25]。SA在H2O2参与下,通过减轻逆境下植物细胞的结构变化、调节抗氧化酶活性和质外体蛋白含量提高植物的抗寒性[26]。刘伟等、常云霞等、王纪忠等的研究表明,SA通过提高低温胁迫下黄瓜幼苗SOD、POD和CAT活性,增强叶片细胞膜的稳定性,缓解低温胁迫造成的冷害[27-29]。本试验中,叶面喷施1 mmol/L SA明显增强了低温处理下黄瓜幼苗的SOD1~SOD4、POD1、POD3~POD6、CAT1和CAT2酶带,表明SA通过影响抗氧化酶同工酶表达参与黄瓜植株低温耐性的调节。
综上所述,叶面喷施2 mmol/L CaCl2明显增强了低温处理下黄瓜幼苗CAT同工酶酶带的强度,叶面喷施1 mmol/L SA明显增强了黄瓜幼苗SOD1~SOD4、POD1、POD3~POD6、CAT1和CAT2酶带,表明钙和SA通过影响抗氧化酶同工酶表达参与黄瓜植株低温耐性的调节。
参考文献:
[1]闫世江,司龙亭,马志国,等. 不同来源黄瓜亲本苗期耐低温鉴定[J]. 沈阳农业大学学报,2006,37(5):766-768.
[2]李益清,李天来. 外源钙及钙抑制剂对番茄耐弱光特性的调控作用[J]. 西北农业学报,2011,20(8):121-126.
[3]张化生,郭晓冬,王 萍. 低温胁迫下硝酸钙处理对辣椒幼苗抗冷性的影响[J]. 甘肃农业大学学报,2008,43(2):66-69.
[4]李美如,刘鸿先,王以柔,等. 水稻幼苗冷锻炼过程中钙的效应[J]. 植物学报,1996,38(9):735-742.
[5]王凤华,王贵学,赖钟雄,等. CaCl2处理对茄子幼苗膜脂过氧化的影响[J]. 江西农业大学学报,2005,27(4):545-547,552.
[6]Ahtúzar-Molina A R,Munoz-Sánchez M,Vázquez-Flota F,et al. Phospholipidic signaling and vanillin production in response to salicylic acid and methyl jasmonate in Capsicum chinense L.cells[J]. Plant Physiology and Biochemistry,2011,49:151-158.
[7]李 亮,董春娟,尚庆茂. 内源水杨酸参与黄瓜叶片光合系统对低温胁迫的响应[J]. 园艺学报,2013,40(3):487-497.
[8]张爱群,沈文梅. 乙醇对超氧化物歧化酶活性邻联〔二〕茴香胺染色法的影响[J]. 生物化学与生物物理进展,1993,20(2):161-162.
[9]王秀芬. 过氧化物酶同工酶最佳染色法[J]. 河北农业大学学报,1990,13(4):78-80.
[10]陈毓荃. 生物化学实验方法和技术[M]. 北京:科学出版社,2002:239-241.
[11]周光宇. 有关同工酶分析的几个问题[J]. 植物生理学通讯,1983(1):1-4,9.
[12]Carpaneto A,Ivashikina N,Levchenko V A,et al. Cold transiently activates calcium-permeable channels in Arabidopsis mesophyll cells[J]. Plant Physiology,2007,143(1):487-494.
[13]Hepler P K. Calcium:a central regulator of plant growth and development[J]. Plant Cell,2005,17(8):2142-2155.
[14]李晓明,陈劲枫,逯明辉,等. 低温下钙对黄瓜幼苗抗氧化酶活性及POD同工酶谱的影响[J]. 西北植物学报,2006,26(2):241-246.
[15]李天来,高晓倩,刘玉凤. 夜间低温胁迫下钙对番茄幼苗根系活力及活性氧代谢的调控作用[J]. 西北农业学报,2011,20(8):127-132.
[16]彭向永,徐永茹,唐 萍,等. 外源钙对低温胁迫下甜瓜幼苗膜稳定性和渗透调节物质的影响[J]. 曲阜师范大学学报:自然科学版,2011,37(1):81-84.
[17]廖金柯,赵 克,胡小燕,等. 低温胁迫后钙对棉花幼苗的影响[J]. 江苏农业科学,2012,40(9):90-91.
[18]王 芳,王丹丹,赵 娟,等. 钙对低温胁迫下玉米幼苗氧化损伤的保护作用[J]. 干旱地区农业研究,2014,32(1):155-160.
[19]姜秀梅,秦 勇,郭光照,等. 外源物质处理对辣椒幼苗抗冷性的影响[J]. 中国农学通报,2013,29(25):87-92.
[20]李兆亮,原永兵,刘成连,等. 黄瓜细胞中水杨酸的信号传递研究[J]. 植物学报,1998,40(5):45-51.
[21]Popova L P,Maslenkova L T,Yornanova R Y,et al. Expogenous treatment with salicylic acid attenuates cadmium toxicity in pea seedlings[J]. Plant Physiology and Biochemistry,2009,47:224-231.
[22]Saidi I,Ayouni M,Dhieb A,et al. Oxidative damages induced by short-term exposure to cadmium in bean plants:protective role of salicylic acid[J]. South African Journal of Botany,2013,85:32-38.
[23]Shi Q H,Zhu Z J. Effects of exougenous salicylic acid on manganeses toxicity,element contents and antioxidative system in cucumber[J]. Environmental and Experimental Botany,2008,63:317-326.
[24]Hayat Q,Hayat S,Irfan M,et al. Effect of exogenous salicylic acid under changing environment:a review[J]. Environmental and Experimental Botany,2010,68(1):14-25.
[25]马德华,庞金安,李淑菊,等. 温度逆境锻炼对高温下黄瓜幼苗生理的影响[J]. 园艺学报,1998,25(4):39-44.
[26]孟雪娇,邸 昆,丁国华. 水杨酸在植物体内的生理作用研究进展[J]. 中国农学通报,2010,26(15):207-214.
[27]刘 伟,艾希珍,梁文娟,等. 低温弱光下水杨酸对黄瓜幼苗光合作用及抗氧化酶活性的影响[J]. 应用生态学报,2009,20(2):441-445.
[28]常云霞,徐克东,陈 璨,等. 水杨酸对低温胁迫下黄瓜幼苗叶片抗寒生理指标的影响[J]. 北方园艺,2013(12):1-4.
[29]王纪忠,周 青. 外源水杨酸对持续低温胁迫下黄瓜幼苗生长和细胞膜稳定性的影响[J]. 长江蔬菜,2010(16):29-32.
