豹纹鳃棘鲈消化道粘液细胞的类型与分布
2016-03-06王永波李向民
王永波,张 杰,李向民
(海南省海洋与渔业科学院,海口 571203)
豹纹鳃棘鲈消化道粘液细胞的类型与分布
王永波,张 杰,李向民
(海南省海洋与渔业科学院,海口 571203)
应用AB-PAS染色法对工厂化养殖条件下豹纹鳃棘鲈(Plectropomus leopardus)消化道粘液细胞的类型与分布进行了观察研究。结果显示:豹纹鳃棘鲈粘液细胞在消化道各部位均有分布,不同部位的粘液细胞密度和类型均有差异,豹纹鳃棘鲈消化道粘液细胞可分为Ⅰ型、Ⅱ型、Ⅲ型、Ⅳ型。其中,食道粘液细胞以Ⅲ型为主,胃以Ⅰ型为主,幽门盲囊和肠以Ⅲ型为主,直肠以Ⅲ型和Ⅳ型为主;胃部粘液细胞分布密度最大,达(111.37±21.74)cell·mm-2,极显著高于消化道其它部分,直肠、幽门盲囊、肠的粘液细胞分布密度分别为(35.09±8.74)、(30.23±5.37)、(28.00±4.67)cell·mm-2,相互之间差异不显著,食道的粘液细胞分布密度最低,为(10.83±1.94)cell·mm-2,与消化道其它部分相比差异极显著。另外,消化道各段单位面积内粘液细胞的分泌能力顺序为食道>直肠>肠>幽门盲囊>胃。通过对消化道各部位粘液细胞的分类对比可以发现,粘液细胞的类型和分布与其所在部位的功能有密切关系。
豹纹鳃棘鲈;消化道;粘液细胞
豹纹鳃棘鲈(Plectropomus leopardus),隶属于鲈形目(Perciformes),鮨科(Serranidae),鳃棘鲈属(Plectropomus),俗称东星斑,属暖水性岛礁鱼类,主要分布于西太平洋至印度洋海区,北自日本南部,南到澳洲,东至斐济,西到非洲东岸、红海等地[1]。国外学者对豹纹鳃棘鲈的研究开展较早,多集中在生态学领域[2-3];国内学者在该鱼的人工繁殖[4]、育苗[5]、养殖[6]、消化生理[7-8]、发育生物学[9]、区系分类[10]等领域做了大量研究工作,目前在我国海南、山东已形成从亲鱼培育、育苗、养殖、销售运输等完善的产业链,养殖经济效益显著。
鱼类的粘液细胞能分泌多种活性物质,如粘多糖、糖蛋白、免疫球蛋白及各种水解酶类,对鱼的许多生理功能有重要影响。对消化道粘液细胞进行深入研究,有助于对鱼类生长、发育等基本生理机制的理解,对鱼类的养殖和病害防治也具有重要的意义[11]。对于鱼类粘液细胞的研究,国外开展得较早,已对多种鱼类如虹鳟(Oncorhynchus mykiss)[12]、大西洋鲑(Salmosalar)[13]等的粘液细胞进行了研究,国内有关这方面的研究也较多,如波纹唇鱼(Cheilinus undulates)[14]、驼背鲈(Cromileptes altivelis)[15]、多鳞四指马(Eleutheronema rhadinum)[16]等,但是有关豹纹鳃棘鲈消化道粘液细胞方面的研究还未见报道。本文对工厂化养殖条件下的豹纹鳃棘鲈消化道粘液细胞进行了观察研究,以期为豹纹鳃棘鲈的消化生理学基础研究和规模化养殖生产提供资料。
1 材料与方法
1.1 材料
实验鱼取自海南省海洋与渔业科学院琼海科研基地,该批鱼为2014年在工厂化养殖模式下养成的豹纹鳃棘鲈商品鱼,全程投喂人工配合饲料。2014年10月,随机抽取5 ind平均体长(28.20±1.35)cm、平均体质量(485.50±75.90)g、体表完整、体型正常的豹纹鳃棘鲈成鱼,饥饿48 h,待其消化道中食物及粪便排空后备用。
1.2 方法
将实验鱼麻醉后沿排泄孔前部剪开,打开腹腔,取材部分由前到后分别为食道、胃、幽门盲囊、肠、直肠。切取组织块,用4%多聚甲醛液固定,常规脱水,石蜡包埋,Leica RM 2135型切片机切片,厚度为4μm,AB-PAS染色,中性树胶封片,分别观察统计粘液细胞的类型、数量及大小,并计算粘液细胞的分布密度。
每个组织随机抽取5张切片,每张切片随机取10个视野,分别测量每一视野中0.01 mm2范围内各类型粘液细胞的总数,作为该种粘液细胞的密度;测量细胞的长径和短径来表示粘液细胞的大小;以消化道各部分粘液细胞的表面积(1/4π×长径×短径)与其细胞分布密度的乘积表示粘液细胞的相对分泌能力,对消化道各部的分泌能力进行计算并比较。
用Motic BA410显微镜观察,Motic Images Advanced 3.2软件摄影和测量。测得数据用Excel和SPSS 19.0软件进行系统分析。
2 结果与分析
2.1 豹纹鳃棘鲈消化道粘液细胞的类型和分布
参照尹苗等[17]对鱼类粘液细胞类型的划分方法,根据AB-PAS染色结果,将豹纹鳃棘鲈粘液细胞分成4种类型:Ⅰ型为红色,AB阴性,PAS阳性,含中性粘多糖;Ⅱ型为蓝色,AB阳性,PAS阴性,含酸性粘多糖;Ⅲ型为紫红色,AB与PAS均为阳性,主要含有PAS阳性的中性粘多糖,同时含有少量AB阳性的酸性粘多糖;Ⅳ型为蓝紫色,AB与PAS同样均为阳性,但主要含有AB阳性的酸性粘多糖,同时含有少量PAS阳性的中性粘多糖。
豹纹鳃棘鲈消化道各部粘膜层上皮中都有粘液细胞分布,但各部粘液细胞的类型和密度存在差异。
食道粘膜层向食道管腔突出,形成指状突起的纵形褶皱,每个纵褶又发出若干个指状突起的二级褶皱,形成食道绒毛,绒毛顶端部分未见有粘膜细胞,绒毛中下端有大量的粘液细胞。粘膜层以Ⅲ型粘液细胞为主,并含有Ⅰ型和Ⅳ型粘液细胞,Ⅱ型粘液细胞为极少量。Ⅲ型粘液细胞在粘膜层均有分布,Ⅰ型粘液细胞多分布在绒毛基部,Ⅱ型和Ⅳ型粘液细胞分布在绒毛中上部及游离部,数量较少(图1-1,图1-2)。可见,在食道的粘膜层中,粘液细胞所含物质是以含中性粘多糖为主,并有少量酸性粘多糖的混合性粘液物质。
胃部的粘液细胞,与关海红等[18]的研究相类似,含有两种粘液细胞,其中胃粘膜上皮表面的粘液细胞,AB-PAS染色呈红色,即Ⅰ型粘液细胞,胃粘膜的基底部的胃腺细胞,AB-PAS染色呈紫红色,即Ⅲ型粘液细胞(图1-3,图1-4)。可见,在胃的粘膜层上皮,粘液细胞所含物质以中性粘多糖为主;在胃粘膜的基底部,粘液细胞所含物质是以含中性粘多糖为主的混合性粘液物质。
幽门盲囊和肠相似,粘膜层以Ⅲ型粘液细胞为主,并含有少量的Ⅰ型粘液细胞。Ⅲ型粘液细胞在柳叶状皱褶上均有分布,在相邻皱褶的隐窝处密度更大,Ⅰ型粘液细胞主要分布在粘膜皱褶底部(图1-5,图1-6,图1-7,图1-8)。可见,在幽门盲囊和肠的粘膜层,粘液细胞所含物质主要是以含中性粘多糖为主的混合性粘液物质。
直肠粘膜层以Ⅲ型粘液细胞和Ⅳ型粘液细胞为主,并含有少量的Ⅰ型粘液细胞和Ⅱ型粘液细胞。Ⅲ型粘液细胞和Ⅳ型粘液细胞在粘膜层上均有分布,Ⅰ型粘液细胞主要分布在粘膜皱褶底部,少量的Ⅱ型粘液细胞分布在Ⅲ型和Ⅳ型粘液细胞之间(图1-9,图1-10)。可见,在直肠的粘膜层上,粘液细胞所含物质主要是中性粘多糖和酸性粘多糖混合粘液物质。

图1 豹纹鳃棘鲈消化道粘液细胞的显微观察(AB-PAS染色)Fig.1 Light microscopical observation of mucous cells in digestive tract of Plectropomus leopardus(Alcian blue-periodic acid sthiff)
2.2 豹纹鳃棘鲈粘液细胞在消化道各部分的密度及细胞大小
粘液细胞在豹纹鳃棘鲈消化道各部分粘膜层的分布密度见表1。胃的粘膜层粘液细胞分布密度最大,为(111.37±21.74)cell·mm-2,极显著高于消化道其它部分;直肠、幽门盲囊、肠的粘液细胞分布密度分别为(35.09±8.74)、(30.23 ±5.37)、(28.00±4.67)cell·mm-2,差异不显著;食道的粘液细胞分布密度最低,为(10.83± 1.94)cell·mm-2,与消化道其它部分相比,差异极显著。
豹纹鳃棘鲈消化道各部分粘液细胞的大小不一,存在显著性差异(表2)。食道部位粘液细胞的平均长径最长,为(22.24±8.88)μm,与消化道其它部分相比,差异极显著;其次为直肠,其粘液细胞的平均长径为(13.29±4.71)μm,与消化道其它部分相比,差异也极显著;幽门盲囊和肠的粘液细胞平均长径分别为(9.11±2.10)μm和(8.59±2.63)μm,它们之间的差异不显著,但与消化道其它部分相比差异极显著;胃的粘液细胞的平均长径最短,为(5.11±1.49)μm,与消化道其它部分相比差异极显著。食道部位粘液细胞的平均短径最长,为(14.66±6.09)μm,与消化道其它部分相比差异极显著;其次为直肠,其粘液细胞的平均短径为(7.84±2.43)μm,与消化道其它部分相比差异也极显著;肠和幽门盲囊的粘液细胞平均短径分别为(5.32±2.02)μm和(5.32±1.56)μm,它们之间的差异不显著,但与消化道其它部分相比差异极显著;胃的粘液细胞的平均短径最短,为(2.56±0.90)μm,与消化道其它部分相比差异极显著。
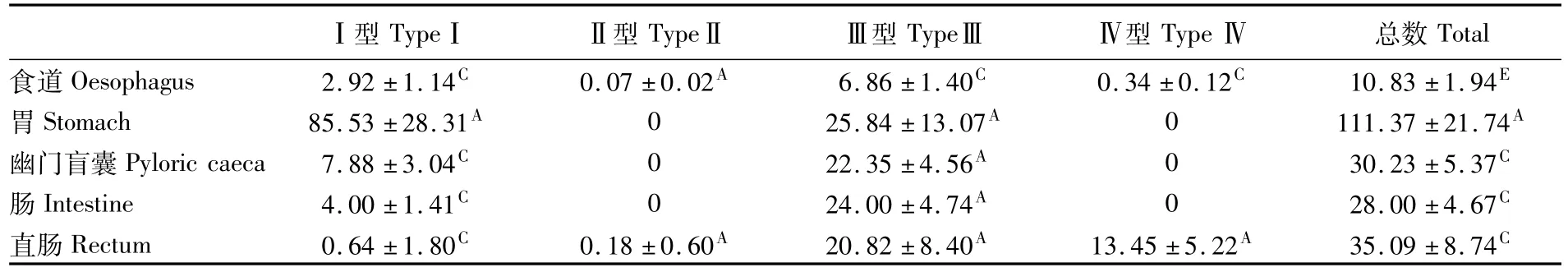
表1 豹纹鳃棘鲈消化道粘液细胞的主要类型和分布密度Tab.1 The density of mucous cells in the digestive tract of Plectropomus leopardus(cell·mm-2,mean±SD)
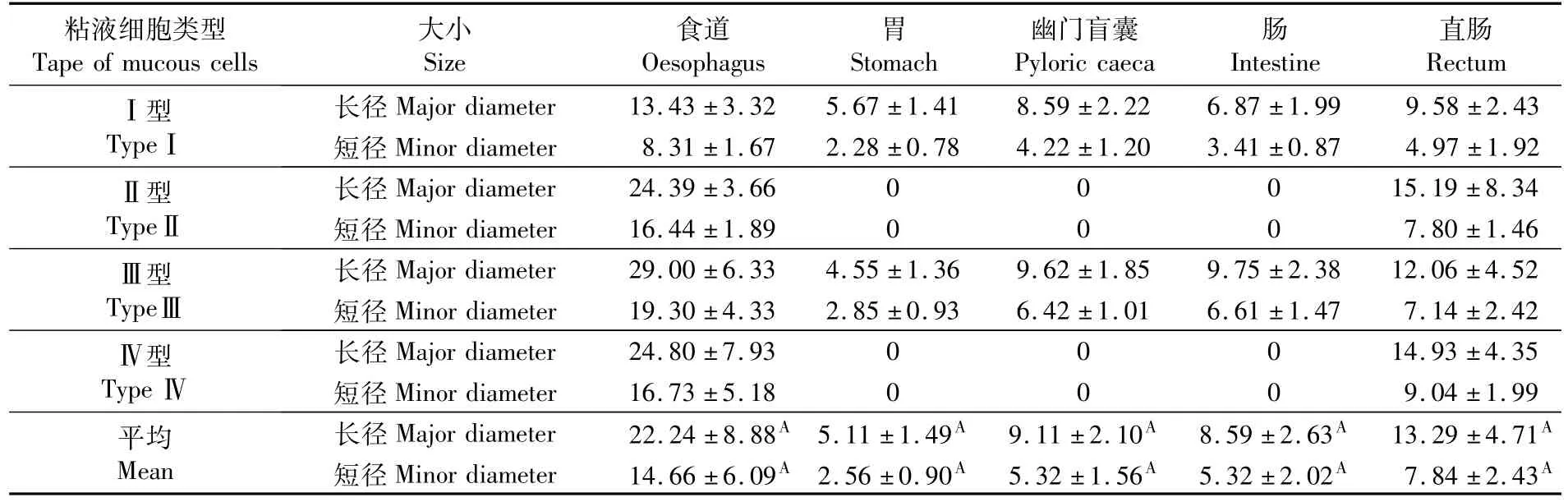
表2 豹纹鳃棘鲈消化道不同部位粘液细胞的类型和大小Tab.2 Tape and size of mucous cells in different parts of the digestive tract of Plectropomus leopardus(μm,mean±SD)
食道粘膜层中的粘液细胞密度虽小,但其体积很大,位于绒毛中下端的多呈圆球型,基部的多呈梨状或囊状;胃部粘膜上皮的粘液细胞多呈单层柱状,胃粘膜的基底部多为圆球型;幽门盲囊和肠的粘液细胞相似,多呈圆形和杯状,粘膜层基底处多呈囊状;直肠的粘液细胞密度和大小相比幽门盲囊和肠的更大,多呈圆形和杯状,粘膜层基底处多呈囊状。
2.3 豹纹鳃棘鲈消化道不同部位粘液细胞分泌能力的比较
豹纹鳃棘鲈消化道各部分的粘液细胞密度、大小都有差异,以消化道各部位的粘液细胞相对总面积(单位面积的细胞数量与细胞平均面积之积)表示粘液细胞的分泌能力P,对消化道各部位的分泌能力进行比较,结果见表3。可知食道和直肠的粘液细胞分泌能力P值分别为(3 485.56±644.21)和(3 044.84±781.01),食道的粘液细胞分泌能力最强,与直肠相比差异显著,与消化道其它部分相比差异极显著;肠、幽门盲囊以及胃的粘液细胞分泌能力P分别为(1 333.68±245.02)、(1 326.51±237.08)和(1 141.62±217.33),它们之间的差异不显著,但与食道和直肠的差异极显著。
3 讨论
3.1 豹纹鳃棘鲈粘液细胞的分类及其在消化道中的分布
本结果表明,豹纹鳃棘鲈胃部粘液细胞的密度最大。由于粘液细胞最初被称为杯状细胞,人们常以它的形态来确定粘液细胞,而豹纹鳃棘鲈胃部粘液细胞不是典型的杯状,从AB-PAS染色结果看,豹纹鳃棘鲈胃部含有粘液细胞。这与花鲈(Lateolabrax japonicus)[19]、哲罗鱼(Hucho taimen)[18]的研究结果相似。豹纹鳃棘鲈胃部含有两种粘液细胞:胃粘膜上皮为表面粘液细胞,单层柱状,AB-PAS染色为红色,属Ⅰ型粘液细胞,含中性粘多糖,具有调节胃中pH值的功能,中性粘多糖在消化道粘膜表面形成一层粘液膜,防止受到酸液的破坏,保护消化道上皮,而中性粘液细胞常与碱性磷酸酶共存,故又有消化功能;胃粘膜的基底部为胃腺细胞,大多为圆形,细胞平均直径比表面粘液细胞的大,AB-PAS染色主要为紫红色,属Ⅲ型粘液细胞,含中性粘多糖为主的混合性粘液物质。胃腺多而发达,具有制造盐酸和胃蛋白酶原的功能,可以消化食物和动物蛋白。
豹纹鳃棘鲈食道的粘液细胞体积较胃、肠的粘液细胞大,这与哲罗鱼相似。粘液细胞在粘膜上皮中的分布形状不同,新形成的囊状细胞与基底膜相连,形成时间长的位于粘膜上皮表层。食道以Ⅲ型粘液细胞为主,并含有Ⅰ型和Ⅳ型粘液细胞,因为食物在酸性的条件进行消化,这种中性偏酸性的粘液环境可能为豹纹鳃棘鲈食道具有消化功能提供证据,同时酸性粘多糖可以利用其酸性成分有效地防止病原菌的侵入。
豹纹鳃棘鲈幽门盲囊与肠的粘液细胞分布密度有一致性,主要为Ⅲ型和Ⅰ型粘液细胞,以中性粘多糖为主的粘液环境起消化作用,少量的酸性粘多糖对消化道上皮起保护作用。直肠的粘液细胞数目也较多,以Ⅲ型和Ⅳ型粘液细胞为主,说明在直肠中中性粘多糖和酸性粘多糖都很丰富。在肠道中,粘液细胞的大小和分布密度从前到后是递增的。
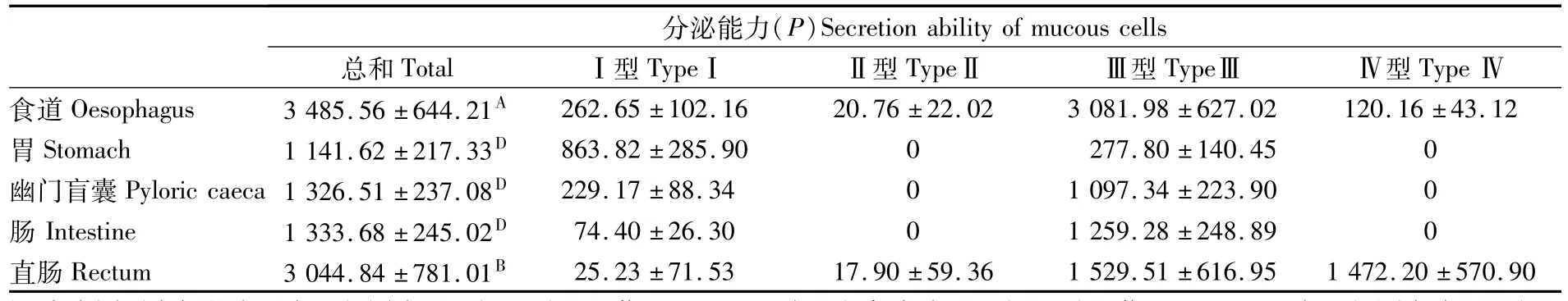
表3 豹纹鳃棘鲈消化道不同部位粘液细胞分泌能力(P)值的比较(mean±SD)Tab.3 Secretion ability of mucous cells in different parts of the digestive tract of Plectropomus leopardus
鱼类消化道粘液细胞的形态、分布密度以及粘液的化学组成不仅与鱼类的种类有关,也与鱼类的生活环境及其食性有关[20],一般说来,草食性的鱼类粘液细胞数量最大,杂食性鱼类粘液细胞的数量居中,肉食性鱼类的粘液细胞数量最少[21]。作为肉食性珊瑚礁鱼类,豹纹鳃棘鲈消化道各段粘液细胞的数量均少于杂食性鱼类如重口裂腹鱼(Schizothorax davidi)[22]、黄姑鱼(Nibea albiflora)[23]、黄鳍鲷(Sparus latus)[24]、长鳍篮子鱼(Siganus oramin,又称黄斑篮子鱼)[25]等,与上述规律一致。
3.2 豹纹鳃棘鲈的消化生理与鱼类消化道各部分泌能力的比较
SINHA[26]研究表明,消化管内壁的粘液在食物通过时有润滑作用,可以防止食物对消化管上皮的机械损伤,并且粘液中含有各种消化酶类,对营养物质的消化起积极作用。不同消化道各部分的分泌能力有差异。粘液细胞在消化道不同部位的分泌能力与对应部位的消化功能相关[27]。
本研究结果表明,豹纹鳃棘鲈食道的粘液细胞分泌能力最强。由于豹纹鳃棘鲈属肉食性鱼类,食物大而复杂,粘液物质多有利于食物顺畅地转移到胃中。另外消化道各部位分泌的酸性粘液物质不仅能润滑食物,还可以软化坚硬的食物,从而更好地保护消化道粘膜层,酸性粘液物质还可以与蛋白酶形成复合物,有稳定酶的作用[28]。在肠道中,粘液细胞的粘液分泌能力从前到后递增,直肠的粘液细胞分泌能力在肠道中最强,这可能与直肠的生理功能有密切关系,直肠与肛门相连,细菌等病原体易侵入,粘液中所含有的免疫性物质可有效除去病原体,同时,直肠中存在大量粘液,有利于粪便的形成和排出,另外直肠也具备食物消化的能力。肠、幽门盲囊以及胃的粘液细胞分泌能力的差异不显著,幽门盲囊与肠的粘液细胞分泌能力的相似性也证明了幽门盲囊可以是肠的分支,可扩大肠对食物的消化吸收面积。又因为幽门盲囊和肠紧连在胃之后,幽门盲囊和肠便于消化从胃中过来的食物,担负着部分消化的功能,这些结果同褐牙鲆的研究相似。胃腺是粘膜上皮的主要组成部分,胃腺细胞既能分泌胃蛋白酶原,同时也能产生盐酸。胃的主要功能之一是消化食物中的蛋白质,发达的胃使食物在胃中停留时间延长,食物被充分地消化,具备发达的胃可以相对容易地将较大的食物进行搅拌和处理成糜状物。食道与直肠会直接与海水接触,与病原生物接触的机会更多,此两部分粘膜层组织上粘液细胞的分泌能力与消化道其它部位相比更强。以上表明消化道各部分粘液细胞分泌能力的强弱是与豹纹鳃棘鲈的消化生理特点相适应的。
[1] LIGHT P R,JONES G P.Habitat preference in newly settled coral trout(Plectropomus leopardus)[J].Coral Reefs,1997,16(2):117-126.
[2] ZHU Z Y,YUE G H.The complete mitochondrial genome of red grouperPlectropomus leopardusand its applications in identification of grouper species[J].Aquaculture,2008,276(1-4):44-49.
[3] KENZO Y,KAZUHISA Y,KIMIO A,et al.Influence of light intensity on feeding,growth,and early survival of leopard coral grouper(Plectropomus leopardus)larvae under mass-scale rearing conditions[J].Aquaculture,2008,279(1-4),55-62.
[4] 王永波,陈国华,林 彬,等.豹纹鳃棘鲈胚胎发育的初步观察[J].海洋科学,2009,33(3):21-26.
WANG Y B,CHEN G H,LIN B,et al.Artificially induced spawning and embryonic development observation of thePlectropomus leopardusLacépnède[J].Marine Sciences,2009,33(3):21-26.
[5] 刘天密,王永波,符书源,等.豹纹鳃棘鲈室外大型水泥池的人工育苗技术[J].海洋渔业,2012,34(4):400-405.
LIU T M,WANG Y B,FU S Y,et al.On the artifical seedling ofPlectropomus leopardusin large outdoor cement pools[J].Marine Fisheries,2012,34(4):400-405.
[6] 王永波,郑 飞,刘金叶,等.豹纹鳃棘鲈工厂化养殖试验[J].热带生物学报,2014,5(1):15-19.
WANG Y B,ZHENG F,LIU JY,et al.Industrial aquaculture experiment of Plectropomus leopardus[J].Journal of Tropical Biology,2014,5(1):15-19.
[7] 张 杰,王永波,李向民,等.工厂化养殖条件下豹纹鳃棘鲈消化系统组织学的观察[J].海洋渔业,2015,37(3):233-243.
ZHANG J,WANG Y B,LIX M,et al.Histological observation on digestive system ofPlectropomus leopardusunder industrialized culture[J].Marine Fisheries,2015,37(3):233-243.
[8] 周邦维,李 勇,高婷婷,等.主要营养素源对工业化养殖豹纹鳃棘鲈生长、体色和消化吸收的影响[J].动物营养学报,2014(5):1387-1401.
ZHOU BW,LIY,GAO T T,etal.Effects ofmain nutrient element and source on growth,body color,digestion and absorption of Plectropomus leopardus in industrialized culture[J].Chinese Journal of Animal Nutrition,2014(5):1387-1401.
[9] MENG Q,DING S,XU X,et al.Ontogenetic development of the digestive system and growth in coral trout(Plectropomus leopardus)[J].Aquaculture,2012,s 334-337(1):132-141.
[10] ZHU Z Y,YUE G H.The complete mitochondrial genome of red grouperPlectropomus leopardusand its applications in identification of grouper species[J].Aquaculture,2008,276(1-4):44-49.
[11] 杨桂文,安利国.鱼类粘液细胞研究进展[J].水产学报,1999,23(4):403-408.
YANG GW,AN L G.A review on the hes of fish mucous cells[J].Journal of Fisheries of China,1999,23(4):403-408.
[12] KHOJASTEH S M B,SHEIKHZADEH F,MOHAMMADNEJAD D,et al.Histological,histochemical and ultrastructural study of the intestine of rainbow trout(Oncorhynchus mykiss)[J].World Applied Sciences Journal,2009,6,(11).
[13] FIRTH K J,JOHNSON S C,ROSS N W.Characterization of proteases in the skin mucus of Atlantic salmon(Salmo salar)infected with the salmon louse(Lepeophtheirus salmonis)and in whole-body louse homogenate.[J].Journal of Parasitology,2000,86(6):1199-1205.
[14] 王永波,陈国华,王 珺,等.波纹唇鱼消化道黏液细胞的类型与分布[J].渔业科学进展,2010,31(5):22-28.
WANG Y B,CHEN G H,WANG J,et al.Types and distribution of mucous cells in the digestive tracts ofCheilinus undulatesRüppell[J].Marine Fisheries Research,2010,31(5):22-28.
[15] 区又君,勾效伟,李加儿.驼背鲈消化系统组织学与组织化学研究[J].海洋渔业,2011,33(3):289-296.
OU Y J,GOU X W,LI J E.Histological and histochemical studies on digestive system ofCromileptes altivelis[J].Marine Fisheries,2011,33(3):289-296.
[16] 黄桂云,张 涛,赵 峰,等.多鳞四指马幼鱼消化道形态学和组织学的初步观察[J].海洋渔业,2012,34(2):154-162.
HUANG G Y,ZHANG T,ZHAO F,et al.On morphology and histology of the digestive tract in young Eleutheronema rhadinum[J].Marine Fisheries,2012,34(2):154-162.
[17] 尹 苗,安利国,杨桂文,等.胡子鲶粘液细胞类型及其在消化道中的分布[J].动物学报,2001,47(专刊):116-119.
YIN M,AN L G,YANG GW,et al.Study on the type of mucous cells and its distribution in alimentary tract ofclarias fuscus[J].Acta Zoologica Sinica,2001,47(s1):116-119.
[18] 关海红,尹家胜.哲罗鱼消化道中黏液细胞的发生和分布[J].水产学杂志,2013,26(5):21-25.
GUAN H H,YIN J S.Ontogenesis and profile of mucilage cells in alimentary canal of taimen(Hucho taimen)[J].Chinese Journal of Fisheries,2013,26(5):21-25.
[19] 谢湘筠,林淑慧,林树根.花鲈消化道粘液细胞的类型及分布[J].福建农业学报,2007,22(3):271-275.
XIE X J,LIN SH,LIN SG.Types and distribution of mucous cells in the digestive tracts ofLateolabraxjaponicus[J].Fujian Journal of Agricultural Sciences,2007,22(3):271-275.
[20] GONA O.Mucous glycoproteins of teleostean fish:a comparative histochemical study[J].Histochemical Journal,1979,11(6):709-718.
[21] SINHA G M.Functional histology of the different regions of the esophagus of a freshwater major carpLabeo calbasuduring the different life history stages[J].Zoologische Beiträge,1977(23):353-360.
[22] 何 敏,张 宇,方 静.重口裂腹鱼消化道黏液细胞类型及分布研究[J].淡水渔业,2007,37(2):24-26.
HE M,ZHANG Y,FANG J.The Study of the mucous cells in the digestive ofSchizothorax davidi[J].Freshwater Fisheries,2007,37(2):24-26.
[23] 辛 俭,薛宝贵,楼 宝,等.黄姑鱼消化道粘液细胞的类型和分布[J].浙江海洋学院学报:自然科学版,2013,32(1):10-14.
YIN J,XUE B G,LOU B,et al.The type and distribution of mucous cells in the digestive tract ofNibea albiflora[J].Journal of Zhejiang Ocean University(Natural Science),2013,32(1):10-14.
[24] 王永翠,李加儿,区又君,等.野生与养殖黄鳍鲷消化道中粘液细胞的类型及分布[J].南方水产科学,2012,8(5):46-51.
WANG Y C,LI J E,OU Y J,et al.Types and distribution of mucous cells in digestive tracts of wild and cultivated yellowfin black porgy(Sparus latus)[J].South China Fisheries Science,2012,8(5):46-51.
[25] 区又君,李加儿,勾效伟,等.黄斑篮子鱼消化道组织学和组织化学研究[J].南方水产科学,2013,9(5):51-57.
OU Y J,LI JE,GOU XW,et al.Histological and histochemical study of digestive tract ofSiganus oramin[J].South China Fisheries Science,2013,9(5):51-57.
[26] SINHA G M.A histochemical study of the mucous cells in the bucco-pharyngeal region of four Indian freshwater fishes in relation to their origin,development,occurrence and probable functions[J].Acta Histochemica,1974,53(2):217-223.
[27] 张伟妮,林 旋,林树根,等.褐牙鲆消化道粘液细胞的类型及分布[J].福建农林大学学报:自然科学版,2009,38(3):280-284.
ZHANGW N,LIN X,LIN SG,et al.Types and distribution of mucous cells in the digestive tract of Paralichthys olivaceus[J].Journal of Fujian Agriculture and Forestry University(Natural Science Edition),2009,38(3):280-284.
[28] 姚荣荣.鳡消化道组织学与免疫组织化学的研究[D].武汉:华中农业大学,2008.
YAO R R.Histological and histochemical study of digestive tract ofElopichthys bambusa[D].Wuhan:Huazhong Agricultural University,2008.
Types and distribution of mucous cells in the digestive tract of Plectropomus leopardus
WANG Yong-bo,ZHANG Jian,LI Xiang-min
(Hainan Academy of Ocean and Fisheries Sciences,Haikou570203)
Plectropomus leopardusis a famous commercial fish species.A complete industrial chain,including cultivation,breeding,sales,transportation,has formed,and the economic benefit is remarkable in Hainan,Shandong and other provinces of China.To carry out the research on mucous cells in the digestive tract will help us understand the basic fish growth and development physiological mechanisms,and also has the vital significance in mariculture and disease control ofPlectropomus leopardus.In this paper,the types and distribution of the digestive tract mucous cells ofPlectropomus leopardusunder factory farming conditions were observed.In October 2014,Plectropomus leoparduswere selected from Qionghai scientific research base of Hainan Academy of Ocean and Fisheries Sciences for the observational study on the fish digestive tract mucous cell types and distribution by using Alcian blue-periodic acid sthiff(AB-PAS).The result shows that mucous cells are distributed in each part of the digestive tract,with different densities and types.Mucous cells in the digestive tract can be divided into typeⅠ,typeⅡ,typeⅢ,and typeⅣ.Among them,the mucous cells of esophagus are mainly typeⅢ;stomach are mainly typeⅠ;pyloric caeca and intestine are mainly typeⅢ;rectum are mainly typeⅢ and Ⅳ.Mucous cells distribution density in stomach(111.37±21.74 cell· mm-2)is the largest and significantly higher than that of other parts in the digestive tract;there is no significant difference among the distribution density in rectum(35.09±8.74 cell·mm-2),pyloric caeca(30.23±5.37 cell·mm-2),and intestine(28.00±4.67 cell·mm-2);the density in esophagus(10.83±1.94 cell·mm-2)is the lowest and significantly different from that of other parts in the digestive tract.For the secretion capacity of mucous cells per unit area in different parts of digestive tract ofPlectropomus leopardus,esophagus and rectum is higher withPvalues of 3485.56±644.21 and 3044.84±781.01 respectively,the secretion capacity of esophagus is the highest and significantly different from that of rectum,and extremely significantly different from other parts in the digestive tract;there is no significant difference among the secretion capacity of intestine(1333.68±245.02),pyloric caeca(1326.51±237.08)and stomach(237.08±217.33).The distribution characteristics and features of the digestive tract mucous cells in different parts of the digestive tract ofPlectropomus leopardusare almost completely consistent with that of carnivorous reef fish,adapted to the digestive physiology characteristic of the fish.
Plectropomus leopardus;digestive tract;mucous cells
S 917
A
1004-2490(2016)05-0478-09
2015-12-25
海南省科学事业费项目(KYYS-2013-18;KYYS-2014-56);海洋公益性行业科研专项(201205028);海南省重大科技项目(ZDZX2013014)
王永波(1983-),男,工程师,主要从事石斑鱼繁养殖研究。E-mail:wangyongbo211@163.com
