‘川早枇杷’头花柱头可授性及花粉活力与花粉管生长的研究
2015-07-05廖明安刘春阳马倩倩杨代宇卢春友周廷国戴盛银
张 潇,廖明安*,何 静,刘春阳,马倩倩,杨代宇,卢春友,周廷国,戴盛银
(1 四川农业大学 园艺学院,四川成都611130;2 汉源县科技局,四川汉源625300;3 汉源县农业局,四川汉源625300)
枇杷(Eriobotryajaponica Lindl.)隶属于蔷薇科(Rosaceae)苹 果 亚 科(Maloideae)枇 杷 属(Eriobotrya)[1]。依成熟期的早晚,枇杷可分为早熟种、中熟种和晚熟种[2],其中早熟种成熟期早,错季节上市,经济效益最高。目前,中国主栽的早熟枇杷品种为‘早钟6号’,是以‘解放钟’为母本,日本特早熟枇杷品种‘森尾早生’为父本进行有性杂交而育成的[3]。‘川早枇杷’属‘早钟6号’枇杷自然芽变,其芽变变异性状稳定,早熟性及果实品质明显优于原母树‘早钟6号’[4]。枇杷开花分三批,‘川早枇杷’第一批花(头花)一般在7~8月开放,花量大,头花果成熟期特早,在12月底可陆续上市,由于正值春节前后,经济效益十分可观,但其头花果与二花果相比,坐果率低成为其生产上的重要限制因子[5]。柱头具有可授性与花粉具有活力是有性生殖器发育成熟的重要标志[6],而花粉在柱头上萌发、花粉管在花柱中生长是受精坐果前非常重要的环节。Ruiz等[7]对43个杏(Prunus armeniaca L.)品种的开花特性展开研究,发现开花时间、花粉萌发率、柱头和花药平面的高度差等开花生物学因子与其最终坐果率高度相关。Abdelgadir等[8]对麻风树(Jatropha curcas L.)花粉活力、花粉萌发和花粉管生长展开研究,认为花粉活力和花粉转移的效率能在一定程度上决定麻风树的繁殖成功率。Bock等[9]通过27年记录26个水仙(Narcissus)品种始花期的时间,发现开花时间和气候变化关系密切,而温度是主要影响因子。枇杷的坐果率与柱头可授性、花粉活力、花粉管动力以及花期温度等密切相关[10-11],但有关早熟枇杷头花授粉受精方面的相关研究还未见报道。为此本研究以特早熟枇杷新品系‘川早枇杷’为试材,对其头花的开花期、柱头可授性、花粉活力、花粉管生长及花期温度等进行了系统研究,以期为揭示其坐果不良机理与调控花果提供理论和实践依据。
1 材料和方法
1.1 材 料
试验于2014年6月~12月在雅安市汉源县唐家乡小关村枇杷园(102°37’E,29°24’N)进行。该区域属亚热带旱暖河谷季风气候,年均气温17.7 ℃,年均 日 照1 496.3h,年 均 降 水 量745.1 mm,年均蒸发量1 248.2 mm,年均无霜期296d。供试材料为树势及栽培管理一致的6年生‘川早枇杷’树20株。
1.2 方 法
1.2.1 开花期、开花持续天数及花序和花器性状观察 通过定株观察‘川早枇杷’头花的开花期,以枇杷全树开花5%~25%为初花期,25%~75%为盛花期,全树开花75%以上至开花基本结束为终花期,并观察记录群株、单株、单序和单花的开花持续天数;于盛花期,随机取花序30个,用游标卡尺测量并记录每个花序主梗长粗、花序的长宽,同时记录花序支轴数、每序花朵总数和花序顶部花数;分别随机取30个花蕾和30朵盛开的花,用游标卡尺测定花蕾长宽、花冠直径、花柱长度,并记录花药数量。
1.2.2 头花花期果园气温的观测 从头花初花期开始至终花期结束为止,采用RC-4HA 型自动温湿度记录仪每隔15min自动采集1次果园的气温(距地面1.5m),从中观察每天的最高和最低温度,并以每天2:00、8:00、14:00和20:00四个时间温度的平均值作为当天的日平均温度。
1.2.3 柱头颜色、黏液分泌观察及柱头可授性检测 于头花盛花期,挂牌标记大蕾期花为参照,选取160朵大蕾期花标记,分别于开花当天(0d)至花后7d,每天随机取20朵标记的花迅速带回实验室,其中10朵用于柱头颜色变化和黏液分泌情况的观察;另10朵花,参照红雨等[11]的方法进行柱头可授性检测,可授性的强弱主要以花柱浸入联苯胺-过氧化氢混合液后气泡的多少及其边缘的颜色为依据进行评价,即气泡越多,颜色越深表示柱头可授性越强,反之亦然,并选取有代表性的材料拍照。
1.2.4 花药颜色及花粉活力观察 于头花盛花期,挂牌标记大蕾期花为参照,选取80 朵大蕾期花标记,分别于从开花当天(0d)至开花后7d,每天各采集花10朵。剥取花药,观察记录花药颜色,并收集花药,花粉活力(率)的测定参照杨芩等[12]的方法进行。
花粉生活力(率)=(变红色花粉数/总花粉数)×100% 。
1.2.5 自花授粉后花粉管生长的荧光显微观察 选择发育良好的花序,于晴天上午去雄后进行‘川早枇杷’自花授粉,授粉前用TTC 法测定花粉活力,共授粉110朵,授粉后用硫酸纸袋套袋隔离并挂牌标记,分别在授粉后2、4、6、8、12、24、36、48、72、96与120h,各取10朵花,参照张潇等[13]的方法固定软化花柱并进行染色,在OLYMPUS-BX51型荧光显微镜下压片观察,并选取有代表性的材料拍照。
1.3 数据分析
采用DPS 系统进行统计分析(Duncan新复极差法进行多重比较)。
2 结果与分析
2.1 ‘川早枇杷’头花开花期、开花持续天数及花序和花器性状
由表1可知,‘川早枇杷’头花花期较早,7月中旬进入初花期,7月下旬至8月上旬为盛花期,8月中旬至下旬为终花期,群株开花持续40d,单株开花持续30d,单序和单花开花持续期分别为17d和8d。由表2可知‘川早枇杷’头花的花序性状,主梗长和粗分别为3.00cm 和0.91cm,花序长和宽分别为13.62 cm 和14.34cm,花序支轴数达10.2个,每序花朵总数多达118.00朵,其中花序顶部花数为18.10朵。由表3可知‘川早枇杷’头花的花器性状,花蕾长和宽分别为1.26cm 和0.69cm,花冠直径为1.69cm,花药数量为20枚,花柱长为0.28cm。

表1 ‘川早枇杷’头花的开花期和开花天数Table 1 Flowering stage and flowering duration of‘Chuanzao loquat’in the first florescence
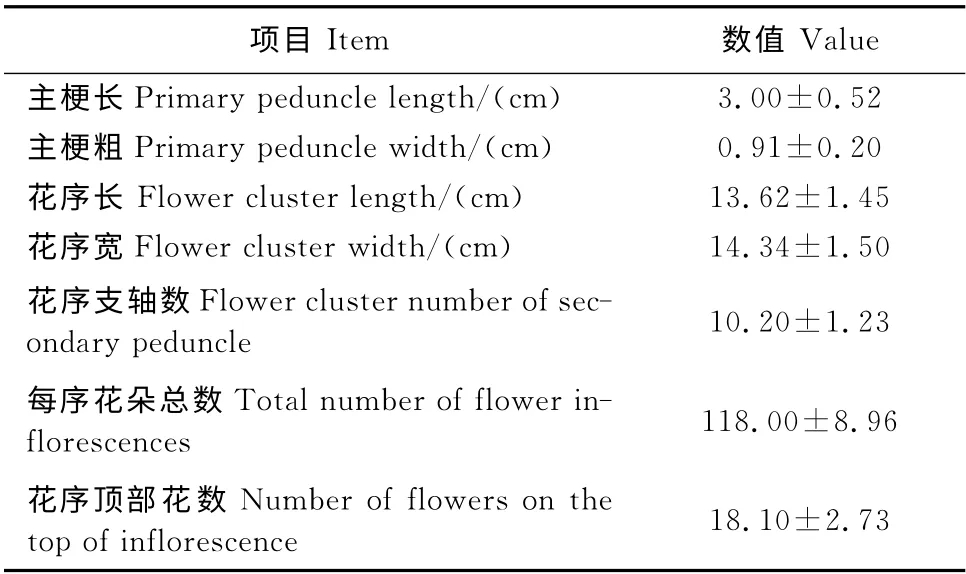
表2 ‘川早枇杷’头花花序性状的观察Table 2 The flower inflorescence characters of‘Chuanzao loquat’in the first florescence
2.2 ‘川早枇杷’头花花期果园气温变化情况
‘川早枇杷’头花花期7月18日~8月27日的果园气温变化情况如图1所示。7月18日~24日为头花初花期,初花期平均温度为27.13℃,日最高温度多出现在中午,初花期平均最高温度达31.35 ℃,而平均最低温度为24.38 ℃。7月28日~8 月9 日为头花盛花期,盛花期平均温度为27.71 ℃,平均最高温度为30.54 ℃,单日最高温度达34.9 ℃,平均最低温度为25.06 ℃。8月13日~27 日为头花终花期,终花期平均温度为23.51℃,平均最高温度27 ℃,平均最低温度21.05 ℃。从整个头花花期的温度来看,初花期和盛花期温度较高,终花期温度相对有所降低。
2.3 ‘川早枇杷’头花柱头可授性和花粉活力
具有生活力的柱头上含有活跃的过氧化物酶,此酶能催化过氧化氢的分解反应,反应释放的氧气量越多酶活性越强,同时在过氧化物酶的作用下,联苯胺被氧化为蓝色或棕色产物,因此气泡越多、颜色越深表示柱头可授性越强。由表4和图2可知,开花当天柱头颜色为浅绿且有黏液分泌,进行柱头可授性检验显示气泡数较多、颜色较深,柱头具有较强的可授性(图2,A);开花后1d柱头周围气泡数最多、可授性最强(图2,B),且黏液分泌最多;而后随着开花时间的延长,柱头黏液分泌逐渐减少,可授性逐渐减弱,开花后2d柱头具有较强可授性(图2,C),开花后3~4d柱头具可授性(图2,D~E),开花后5~6d柱头部分具可授性(图2,F~G),开花后7 d柱头不具有可授性(图2,H)。‘川早枇杷’柱头属于湿柱头型,开花当天柱头表面为浅绿色并布满乳突细胞(图2,I),利于花粉的粘附,但到开花后4d,柱头表面变为褐色,乳突细胞明显减少(图2,J)。相应地,开花当天花药呈嫩黄色,随着花龄的增加,花药颜色由嫩黄变为黄色、黄褐色、褐色、深褐色,最后变为黑色。花粉活力在开花后1d最高(53.17%),其次为开花后2d和开花当天,开花后3~7d,花粉活力随着开花后天数的增加而迅速降低,开花后6~7d花粉活力极低,极显著小于其它各处理。

表3 ‘川早枇杷’头花花器性状的观察Table 3 The flower characters of‘Chuanzao loquat’in the first florescence
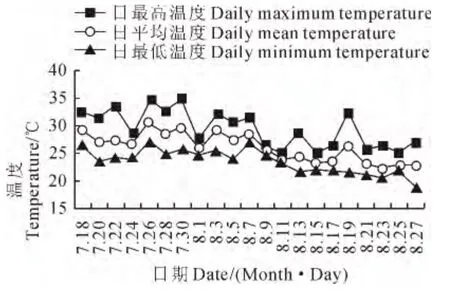
图1 ‘川早枇杷’头花花期果园的日气温动态变化Fig.1 Daily air temperature change curve of the orchard in the first florescence of‘Chuanzao loquat’
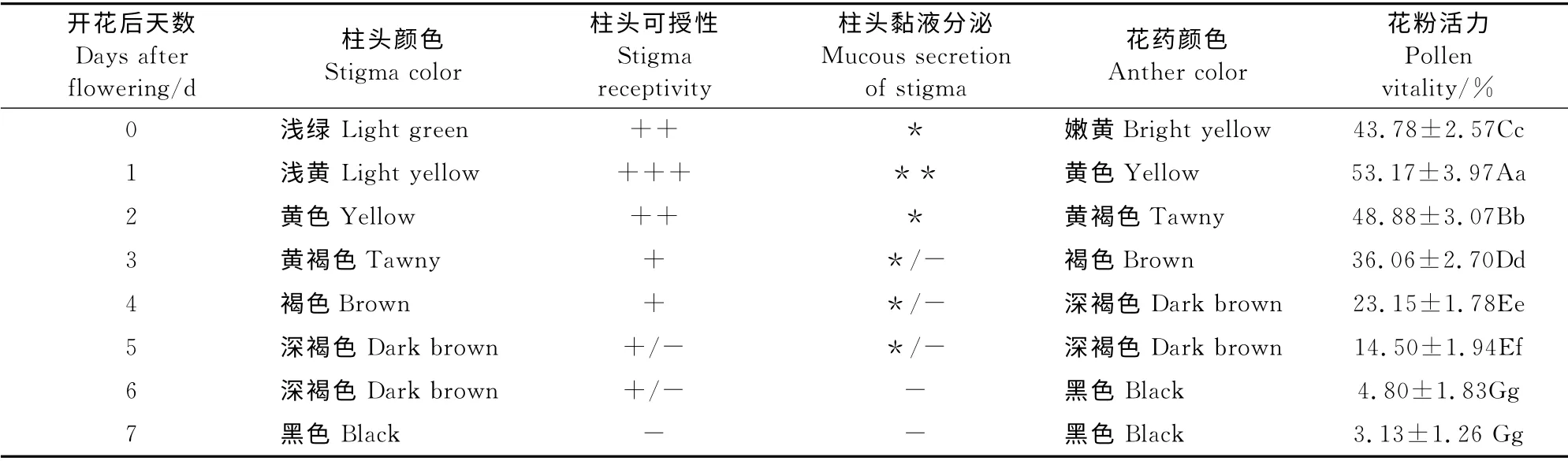
表4 ‘川早枇杷’头花开花后花粉活力及柱头可授性观察Table 4 The stigma receptivity and pollen viability of‘Chuanzao loquat’after flowering in the first florescence
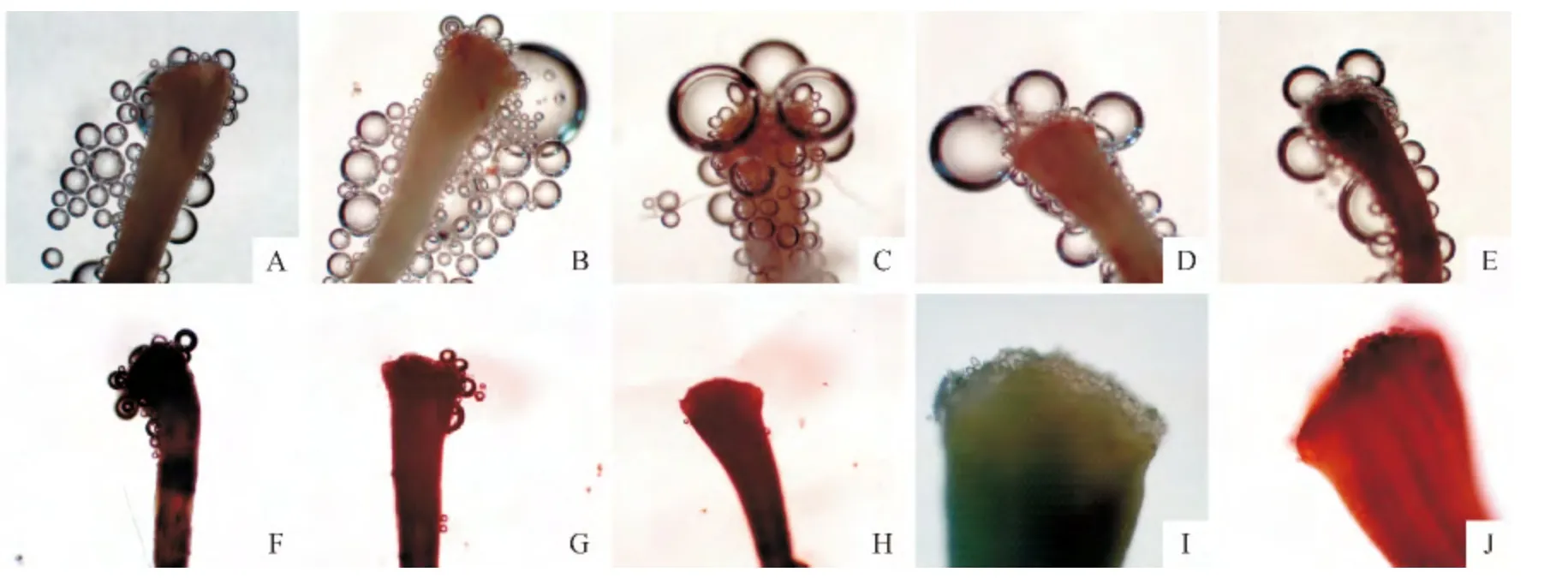
图2 ‘川早枇杷’头花柱头可授性和柱头表面特征观察A.开花当天(0d),柱头具有较强可授性(10×4);B.开花后1d,柱头具有强可授性(10×4);C.开花后2d,柱头具有较强可授性(10×4);D~E.开花后3~4d,柱头具有可授性(10×4);F~G.开花后5~6d,柱头部分具可授性(10×4);H.开花后7d,柱头不具可授性(10×4);I.开花当天(0d),柱头表面布满乳突细胞(10×10);J.开花后4d,柱头乳突细胞较少(10×10)。Fig.2 The stigma receptivity and surface characters of‘Chuanzao loquat’in the first florescence A.Stigma with quite strong receptivity on the flowering day(10×4);B.Stigma with very strong receptivity on 1dafter flowering(10×4);C.Stigma with quite strong receptivity on 2dafter flowering(10×4);D-E.Stigma with receptivity on 3-4d after flowering(10×4);F-G.Stigma with weak receptivity on 5-6dafter flowering(10×4);H.Stigma without receptivity on 7d after flowering(10×4);I.Stigma surface was full of mastoid cells on the flowering day(10×10);J.Stigma with only a few mastoid cells on 4dafter flowering(10×10).
2.4 ‘川早枇杷’头花自花授粉后花粉管生长情况
由图3可知‘川早枇杷’头花自花授粉后花粉管生长情况。自花授粉前用TTC 法测定花粉活力为50.28%,自花授粉后2h,柱头上未见花粉萌发(图3,A);授粉后4h,花粉萌发并已穿过柱头进入花柱,但萌发率较低(图3,B);授粉后12h,花粉管到达花柱中上部约1/5处(图3,C);授粉后24h,花粉管到达花柱中部,并出现花粉管中下部弯曲生长、荧光效应强烈等现象(图3,D);授粉后36h,花粉管到达花柱中下部约3/5处,花粉管弯曲生长并伴随产生花粉管部分位点荧光效应强烈等现象(图3,E);授粉后48h,花粉管成簇状进入花柱中下部大约4/5处,但数量较少,且花粉管部分位点膨大、荧光明亮(图3,F);授粉后48h,花粉管已抵达花柱基部,但是数量极少,大约1~3根,且花粉管顶部膨大,荧光效应强烈(图3,G);授粉后96h,花粉管进入胚珠,且数量极少(图3,H)。
3 讨 论
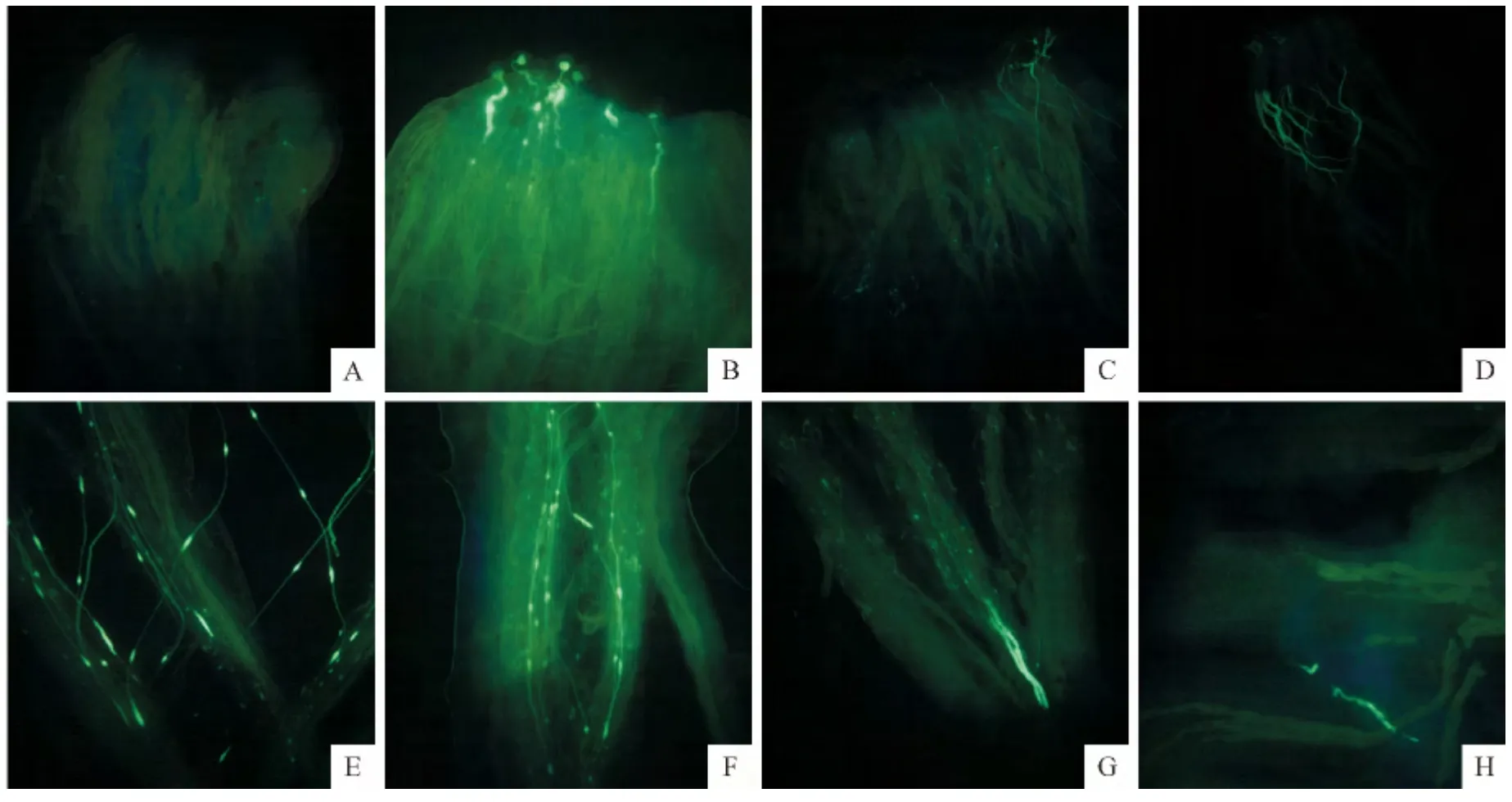
图3 ‘川早枇杷’头花自花授粉后花粉管生长的荧光显微观察A.自花授粉后2h,柱头上未见花粉萌发(10×10);B.自花授粉后4h,花粉已萌发并穿过柱头进入花柱(10×20);C.自花授粉后12h,花粉管到达花柱中上部约1/5处(10×10);D.自花授粉后24h,花粉管到达花柱中部(10×4);E.自花授粉后36h,花粉管到达花柱中下部约3/5处(10×10);F.自花授粉后48h,花粉管进入花柱中下部大约4/5处(10×10);G.自花授粉后48h,极少数的花粉管抵达花柱基部(10×10);H.自花授粉后96h,花粉管抵达胚珠(10×10)。Fig.3 Fluorescence microscopic observation on pollen tube growth of‘Chuanzao loquat’after self-pollination in the first florescence A.Pollens didn not germinate at 2hafter self-pollination(10×10);B.Pollens germinated and pollen tubes through stigma into style at 4hafter self-pollination(10×20);C.Pollen tubes reached about 1/5of style at 12hafter self-pollination(10×10);D.Pollen tubes penetrated into the middle of style at 24hafter self-pollination(10×4);E.Pollen tubes reached about 3/5of style at 36hafter self-pollination(10×10);F.Pollen tubes reached about 4/5of style at 48hafter self-pollination(10×10);G.A few pollen tubes extended into the base of style at 48hafter self-pollination(10×10);H.Pollen tubes entered the ovule at 96hafter self-pollination(10×10).
柱头是雌蕊顶端接受花粉的部位,柱头表面的形态和生理特征影响着花粉能否成功黏附[14]。柱头可授性越高表明柱头接受有活力的花粉和维持花粉萌发的性能越高[15],具有可授性的柱头是花粉的天然培养基,能为花粉黏附、水合萌发及花粉管在柱头上的生长提供物质支撑[16]。‘川早枇杷’头花柱头具有可授性的持续时间较短约为4~5d,可授性最佳的持续时间也较短仅为开花后1d。柱头颜色变化、黏液分泌与柱头可授性有一定的相关性,随着开花后天数的增加,柱头颜色由浅绿到黄色再到黑色,黏液分泌量由少到多再到少,相应地柱头可授性也从弱到强再到最后丧失。王丽娟等[17]研究发现甜樱桃(Cerasus avium)柱头可授性与乳突细胞破裂有关。本研究中,‘川早枇杷’头花开花当天柱头表面乳突细胞多而明显,增加了柱头吸收花粉的表面积,柱头具有较强可授性,开花后4d,柱头颜色变为褐色,乳突细胞明显减少,柱头可授性降低。花粉在柱头上萌发后需穿过柱头才能进入花柱,李楠等[18]研究发现沙田柚(Citrus grandis)花粉原位萌发后通过乳突细胞间隙进入柱头,并沿柱头区薄壁细胞间隙进入花柱。马纪峰等[19]研究发现拟南芥(Arabidopsis thaliana)花粉可以通过释放某些酶分解乳突细胞壁中的纤维素和半纤维素,从而使花粉管穿过乳突细胞壁。本研究发现,授粉后4h已有少量花粉管穿过柱头进入花柱,但关于其具体是如何穿过柱头进入花柱的还有待进一步研究。
花粉是否能在柱头上萌发并穿过柱头,不仅与柱头可授性有关,与花粉活力也密切相关。而花粉活力的大小、持续时间的长短,又受遗传因素的决定和环境因素的影响。郭尚等[20]研究发现西瓜(Citrullus lanatus Mansfeld)花粉萌发的适宜温度为18~38 ℃,在较高温度下形成的花粉生活力强。杨芩等[12]在研究不同花龄下‘大五星’枇杷的花粉活力时发现,开花后2~5d其花粉活力均维持在较高水平(67.75%~72.65%),开花后6~7d,花粉活力也大于40%。但本研究中,‘川早枇杷’头花花粉活力在开花后1d达到最大(53.17%),而后迅速降低,至开花后6~7d,花粉活力极低(<5%),这可能与‘川早枇杷’头花花期正值高温的夏季,而‘大五星’枇杷花期多为温度较低的秋冬季节有关。
具有活力的花粉在柱头表面萌发后,花粉管穿过柱头进入花柱,花粉管在花柱中的生长速度不仅受品种特性、自身营养条件的影响,还受外界温度等的 影 响[21-22]。霍 宏 亮 等[23]研 究 认 为 环 境 温 湿 度 和种质对川榛(Corylus heterophylla var.sutchuanensis)花粉萌发有重要影响,而花粉管生长速度受环境温湿度的影响较大,与种质无关。本研究发现,‘川早枇杷’授粉后48h,只有极少数的花粉管能到达花柱基部,一方面可能是因为自花授粉存在一定的不亲和,能穿过柱头进入花柱和在花柱中生长的花粉管数量都相对较低;另一方面,可能是因为花期高温不利于花粉黏附萌发和花粉管在花柱中的极性生长。此外,‘川早枇杷’头花花期早,与其它品种的枇杷往往花期不遇,所以其多进行自花授粉,加之花期高温天气不利于授粉受精,头花果的种子数平均为2.3粒[5],种子数较少,且其头花果具有极早熟的特性,‘川早枇杷’头花果可以作为培育早熟少核枇杷的一种重要种质资源。综上所述,‘川早枇杷’头花花期高温天气不利于其花粉萌发和花粉管生长,柱头具有最佳可授性的持续期较短、花粉活力降低迅速以及能抵达花柱基部和进入胚珠的花粉管数量少等诸多因素的综合作用是造成其头花坐果不良的重要原因。
[1] ZHENG G H(郑国华),ZHANG H Y(张贺英),ZHONG X R(钟秀容).Changes in cell ultra-structure,membrane permeability and protective enzyme activity in Eriobotryajaponica Lindl.leaves under cold stress[J].Chinese Journal of Eco-Agriculture(中国生态农业学报),2009,17(4):739-745(in Chinese).
[2] LI Z K(李泽坤),ZHOU T(周 婷).Research progress on loquat breeding[J].Xiandai Horticulture(现代园艺),2013,(1):6-8(in Chinese).
[3] LU X M(陆修闽),CHEN J Y(陈菁瑛),ZHANG L M(张丽梅),et al.Observation and comparison on pollen morphology of a new hybrid loquat variety‘Zaozhong 6’and its parents[J].Acta Horticulturae Sinica(园艺学报),2002,29(3):271-273(in Chinese).
[4] MEI L Y,LIAO M A,REN Y J,et al.Study on fruit quality,phonological phases and shoot histomorphology of a new bud mutant line,‘Chuanzao Loquat’[J].Agricultural Science &Technology,2012,13(9):1 881-1 884.
[5] 鲍荣粉.‘川早枇杷’生物学特性调查及疏果套袋对果实品质的影响[D].成都:四川农业大学,2013.
[6] CRAWFORD B C W,YANOFSKY M F.The formation and fuction of the female reproductive tract in flowering plants[J].Current Biology,2008,18(20):927-978.
[7] RUIZ D,EGEA J.Analysis of the variability and correlations of floral biology factors affecting fruit set in apricot in a Mediteranean climate[J].Scientia Horticulturae,2008,115(2):154-163.
[8] ABDELGADIR H AJOHNSON S DVAN STADEN J.Pollen viabilitypollen germination and pollen tube growth in the biofuel seed crop Jatropha curcas(Euphorbiaceae)[J].South African Journal of Botany,2012,79:132-139.
[9] BOCK A,SPARKS T H,ESTRELLA N.Climate sensitivity and variation in first flowering of 26Narcissus cultivars[J/OL].International Journal of Biometeorology(2014-8-26).http://link.springer.com/article/10.1007/s00484-014-0885-6
[10] 杨 芩.枇杷授粉受精生物学研究与S基因克隆[D].成都:四川农业大学,2013.
[11] HONG Y(红 雨),FANG H T(方海涛),NA R(那 仁).Pollen viability and stigma receptivity of Prunus mongolica Maxim.[J].Guihaia(广西植物),2006,26(6):589-591(in Chinese).
[12] YANG Q(杨 芩),FU Y(付 燕),WANG Y Q(王永清),et al.Effects of flower age on stigma receptivity and pollen vitality of loquat[J].South China Fruit(中国南方果树),2012,41(4):91-93(in Chinese).
[13] ZHANG X(张 潇),LIAO M A(廖明安),WANG Y Q(王永清),et al.Research on the difference in the pollen tube growth of a low seediness line of loquat[J].Acta Bot.Boreal.-Occident.sin.(西北植物学报),2014,34(1):26-31(in Chinese).
[14] LI J(李 晶),JIN L(金 樑),DENG ZH G(邓志刚),et al.Pollen-pistil interaction and its molecular basis in angiosperms[J].Pratacultural Science(草业科学),2014,31(1):161-167(in Chinese).
[15] GUO Y(郭 媛),SHAO Y Q(邵有全),GUO B B(郭宝贝),et al.Relationship of pear pollen and stigma development with temperature[J].Chinese Journal of Eco-Agriculture(中国生态农业学报),2014,22(12):1 446-1 452(in Chinese).
[16] JAVIER S,PILAR R,MARIA H.Asynchronous development of stigmatic receptivity in the pear(Pyrus communis;Rosaceae)flower[J].American Journal of Botany,2003,90(1):78-84.
[17] WANG L J(王丽娟),LIU L D(刘林德),ZHANG L(张 莉),et al.Stigma receptivity,stigma morphology and fruit set of Yantai sweet cherry(Cerasus avium)[J].Chinese Bulletin of Botany(植物学报),2011,46(1):44-49(in Chinese).
[18] LI N(李 楠),ZHANG X H(张杏辉),XUE M N(薛妙男),et al.Studies on structure of stigma of Shatinyu and the ways of pollen tube enter the stigma[J].Journal of Guangxi Normal University(广西师范大学学报),1995,13(4):58-63(in Chinese).
[19] MA J F(马纪峰),WANG X L(王秀玲).Ultrastructures of pollen and papillar cell of Arabidopsis during pollination[J].Chinese Journal of Cell Biology(中国细胞生物学学报),2011,33(8):892-896(in Chinese).
[20] GUO SH(郭 尚),WANG X Y(王秀英).Effects of the different factors on vigor power of the watermelon pollen[J].Acta Agriculturae Boreali-sinica(华北农学报),2006,21(3):91-94(in Chinese).
[21] THOMAS D,NONI F.Male-female crosstalk during pollen germination,tube growth and guidance,and double fertilization[J].Molecular Plant,2013,6(4):1 018-1 036.
[22] PASONEN H L,KÄPYLÄM,PULKKINEN P.Effects of temperature and pollination site on pollen performance in Betula pendula Roth-evidence for genotype-environment interactions[J].Theor.Appl.Genet,2000,100:1 108-1 112.
[23] HUO H L(霍宏亮),ZHANG R Q(张日 清),MA Q H(马庆 华),et al.Stigma receptivity and characteristics of pollen tube growth of Corylus kweichowensis[J].Forest Research(林业科学研究),2014,27(3):403-409(in Chinese).
