不同培养基对禾草内生真菌Epichloё生长与产孢的影响
2015-06-24李秀璋郭长辉李春杰
柳 莉,李秀璋,郭长辉,李春杰
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
不同培养基对禾草内生真菌Epichloё生长与产孢的影响
柳 莉,李秀璋,郭长辉,李春杰
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州 730020)
本研究以分离自西藏早熟禾(Poatibetica)、甘肃臭草(Melicaprzewalskvi)和麦宾草(Elymustangutorum)的6个内生真菌菌株(分别为西藏早熟禾内生真菌Ep-003和Ep-004,甘肃臭草内生真菌Em-014和Em-016,麦宾草内生真菌Ee-001和Ee-003)为研究对象,分别在马铃薯葡萄糖琼脂(PDA)、麦秆煎液琼脂(WSA)、玉米粉琼脂(CMA)和水琼脂(WA)4种培养基上培养4周后,观测菌落生长速率、菌落形态特征、产孢量和孢子特性。结果表明,同一菌株在不同培养基上的生长速率一般为PDA和CMA上的最大,WSA和WA上最小;不同菌株在同一培养基上,西藏早熟禾的Ep-003菌株生长最快,在PDA上直径最大,为46.42 mm,麦宾草内生真菌次之,甘肃臭草的Em-014菌株生长最慢,在WA上直径最小,为7.07 mm;在4种培养基上,菌株的产孢量有显著差异(P<0.05),分别为Ep-003只在WA上产孢,其他5种菌株在WA和WSA培养基上产孢较多,在PDA和CMA上产孢较少; Em-016和Ep-004在4种培养基上的孢子长度无显著差异(P>0.05),Em-014在WSA上的孢子长最小,为3.77 μm,其他菌株无显著差异(P>0.05),Ee-003在PDA上孢子长最长,为8.43 μm,在其他3种培养基上无显著差异(P>0.05);Ep-004、Em-016和Ee-003在4种培养基上孢子梗长度均无显著差异(P>0.05),Em-014在WA上孢子梗最长,为16.74 μm,在WSA上最短,为14.09 μm,Ee-001在CMA上孢子梗最长,为18.59 μm,在WSA上最短,为13.40 μm。
共生真菌;分生孢子;生长速度;产孢特征
禾草内生真菌(Endophytic fungi)是在宿主体内度过全部或大部分的生命周期, 而宿主植物不显示任何外部症状的一类真菌[1]。这个类群的内生真菌一般是指Epichloё属的真菌及其无性阶段Neotyphodium属,根据最新国际真菌命名法规,现统一称为Epichloё内生真菌[2]。有关Epichloё属及其无性阶段内生真菌生理学与生物学特性的报道相对较少。有关对Epichloё内生真菌生长条件及其产孢特性影响的研究主要集中在培养基种类[3]、培养温度[4]、糖分含量及最优碳源[5-6]等。培养基的种类对Epichloё属3种内生真菌的生长速度及其菌落形态影响显著[3]。Neotyphodium属的5种内生真菌生长所需最适糖分含量差异显著[5]。此外,White等[6]对产生和不产生子座的E.typhinum在3种不同糖浓度培养基中的菌落生长情况进行了研究,发现糖分含量对其生长影响显著,并筛选出了最适碳源培养基。温度不仅对内生真菌菌落生长速度及菌落形态造成影响[3],同时对内生真菌的产孢特性影响显著[4]。国内关于内生真菌产孢机制的研究主要集中在甘肃内生真菌(E.gansuense)[7]及分离自高羊茅(Festucaarundinacea)[7]、黑麦草(Loliumperenne)[8]和中华羊茅(Festucasinensis)[9]的内生真菌。而对于分离甘肃臭草(Melicaprzeualskyi)、西藏早熟禾(Poatibetica)和麦宾草(Elymustangutorum)的内生真菌的生物学与生理学的研究甚少。
甘肃臭草是广泛分布于甘肃、青海等地的一种多年生根茎类草本植物,家畜往往因其产生的怪异味道而拒绝采食,是我国西北天然草地上比较常见的一种杂草植被物种[10]。研究发现,遭受甘肃臭草入侵的草地会在短期内演变为以其为优势种的退化草地,这种毒杂草的大量滋生、扩展及蔓延,严重影响着家畜及其他野生草食动物对草地的有效利用[11]。目前,有关甘肃臭草内生真菌的研究仅见于Li等[12]对其分离培养的简要描述,但对于其在不同培养基上的生物学及其产孢特征尚未进行研究报道。
西藏早熟禾是一种禾本科早熟禾属的多年生草本植物,分布在海拔2 500~3 200 m的亚高山草甸[13],抗寒,耐湿冷,生活力极强,在高寒草地分布广,参与度小,数量少,但草质柔嫩,叶量丰富,适口性好,是牛、马、羊常均喜食的优良牧草。有关西藏早熟禾内生真菌的研究主要集中在带菌率[14]、简要体外培养特征[12]等,但是相对较为深入的生物学特征尚未见报道。
麦宾草系禾本科披碱草属多年生草本植物,秆单生或呈疏丛型,须根较粗,高100~120 cm,直径可达5.5 mm,具4~5节,基部节稍曲[15]。麦宾草在我国主要分布于青海、西藏、甘肃、内蒙古等大部地区,国外在尼泊尔等地也有分布。有关麦宾草内生真菌的研究主要集中在其带菌率[14],对于其产孢机制的研究尚未见报道。
Epichloё属内生真菌在体外培养中生长极为缓慢,且在常规培养基上一般不易产孢,而孢子是禾草内生真菌生物学研究的一个重要组成部分。本研究分析不同类型培养基对禾草内生真菌产孢特性的影响,旨在为寻找内生真菌的体外培养的最适培养基提供理论依据,同时为内生真菌分类学、系统学的研究提供一定的理论基础。
1 材料与方法
1.1 试验材料
1.1.1 内生真菌 内生真菌菌株分离自西藏早熟禾、甘肃臭草和麦宾草3种宿主植物,由兰州大学草地保护研究所保存。6个菌株分别根据其宿主禾草的属名缩写,依此分别命名为西藏早熟禾内生真菌Ep-003、Ep-004,甘肃臭草内生真菌Em-014、Em-016和麦宾草内生真菌Ee-001、Ee-003。分别转接至马铃薯葡萄糖琼脂培养基(PDA)上,每个菌株3个重复,22 ℃黑暗培养箱(GXZ-380C,浙江)中生长4周,待用。
1.1.2 培养基 参照Li等[16]的方法,制作马铃薯葡萄糖琼脂培养基(PDA)、麦秆煎液琼脂培养基(WSA)、玉米粉琼脂培养基(CMA)和水琼脂培养基(WA)。PDA制作方法:去皮马铃薯200 g,琼脂粉17 g,加蒸馏水至1 000 g。CMA制作方法:玉米粉300 g,琼脂粉17 g,加蒸馏水至1 000 g。WA制作方法:琼脂粉20 g,加蒸馏水至1 000 g。WSA制作方法:麦秆20 g,琼脂粉17 g,加蒸馏水至1 000 g,其中,麦秆先加入1 000 g蒸馏水,放入温度为120 ℃的高温灭菌锅(HG-50,日本)中进行煮沸灭菌20 min,再用双层纱布过滤至加入琼脂粉的三角瓶中,再进行一次高温灭菌,最后将所有灭过菌的培养基倒入直径为9 cm的培养皿中,每皿约倒25 mL,制成平板培养基。
1.2 方法
1.2.1 不同培养基上的菌落生长 参照Li等[16]的方法,将生长于22 ℃黑暗条件下培养4周的内生真菌用直径为4 mm的打孔器纯培养菌落打成菌饼,随后将菌饼分别转接到PDA、WA、WSA和CMA平板培养基中央,每皿1个菌饼。第4周用数显游标卡尺(溯源测量仪器有限公司,湖南)测定菌落直径,每个菌株3个重复。
1.2.2 产孢量的测定 4周试验结束后进行产孢量的测定,将无菌水加入到生长好的内生真菌培养皿中,用接种环轻轻搅动,使孢子充分落下,滴一滴菌悬液至载玻片上,40×10倍光镜条件下观察每个视野中的孢子数,每种培养基3个重复,共观察10个视野,求平均值。
1.2.3 孢子、孢子梗的测定 培养4周以后,用接种针挑取菌丝少许放入滴有一滴无菌水的载玻片上,盖上盖玻片,轻轻按压,然后在矫正过的40×显微镜(每小格2.44 μm)下测定孢子的长、宽,孢子梗的长、宽,记录格数,计算大小。
1.2.4 数据统计分析 所有数据均用Microsoft Excel录入,采用SPSS 17.0 统计分析软件进行差异显著性分析,Ducan法进行多重比较分析。
2 结果与分析
2.1 不同培养基对内生真菌菌落特征的影响
同一菌株在不同培养基上菌落形态存在差异,菌落正面的特征为:Ep-003在PDA上菌落正面稀疏(图1),在CMA、WSA上较稀疏(图1-3、图1-5),在WA上气生菌丝最稀薄(图1-7);在PDA和WSA上菌落背面为白色至褐色(图1-2,图1-6),而在CMA和WA上菌落背面均为白色(图1-4,图1-8)。Ep-004在PDA上正面为白色,致密,棉状,在其他3种培养基上正面为白色,中央突起,气生菌丝稀薄(图2),其中在WA上气生菌丝最稀薄(图2-8)。背面的特征为:在PDA为白色至褐色,在WSA上为深褐色,在CMA和WA上为白色。Em-014在PDA、WSA和WA上菌落正面为白色,致密,在CMA上气生菌丝稀薄;菌落背面特征与Ep-004在4种培养基上的一致。Em-016的菌落特征与Em-014的相同(图3、图4)。Ee-001和Ee-003在PDA上菌落正面稍稀疏,在WSA上菌落正面致密(图5),棉状,气生菌丝稀薄(图5-3、图5-6),在CMA和WA上菌落正面稀疏,气生菌丝稀薄(表1)。
2.2 不同培养基对内生真菌菌落生长的影响
培养4周后,6个菌株中除Ep-004和Ee-001外,其余菌株在PDA和CMA上的菌落直径显著大于在WSA和WA上的菌落直径(P<0.05)(表2)。
对于西藏早熟禾、甘肃臭草、麦宾草3种内生真菌而言,甘肃臭草的内生真菌菌落直径最小;西藏早熟禾的Ep-003菌株直径最大。对于同一种菌的不同菌株而言,菌落直径也存在差异,如Ep-003在WSA上的菌落直径显著大于Ep-004的(P<0.05),Em-014和Em-016在CMA和WSA上菌落直径差异不显著(P>0.05)(表2)。
2.3 不同培养基对内生真菌产孢特性的影响
除Ep-003只在WA产孢外,其他5个菌株在4种培养基上均产孢(表3)。麦宾草内生真菌在4种培养基上产孢量最高,甘肃臭草内生真菌次之,西藏早熟禾内生真菌的产孢量是3个菌里最少的;Ep-004在4种培养基上都产孢,且在CMA和WSA上产孢较多;甘肃臭草内生真菌在WSA上产孢较多;麦宾草内生真菌在CMA、WSA和WA上产孢较多,其中Ee-001在WA上产孢最多。
对4种培养基上的6个菌株而言,产孢量大小顺序为WSA>WA>CMA>PDA,原因很可能是WSA和WA营养贫乏,适合生殖生长。
在PDA上,菌株Ee-003的孢子最长,其他菌株孢子长度不存在显著差异(P>0.05);CMA上,Ep-004的孢子最长;WSA上,Em-014的孢子最短(表4)。同一菌株在不同培养基上的孢子长度也存在差异,Ep-004在WA上孢子最短;Em-014、Em-016和Ee-001都在WSA上孢子最短;Ee-003在PDA上的孢子最长(表4)。
各菌株的孢子宽在不同培养基上存在一定的差异,其中Ee-003的孢子最宽(表5)。 对不同菌的内生真菌而言,孢子宽度大小为麦宾草>西藏早熟禾>甘肃臭草;对不同菌株而言,甘肃臭草内生真菌的两个菌株在WSA上的宽度均小于在其他3种培养基上的孢子宽;Ee-001在CMA上的孢子宽小于在其他3种培养基上的孢子宽(表5)。
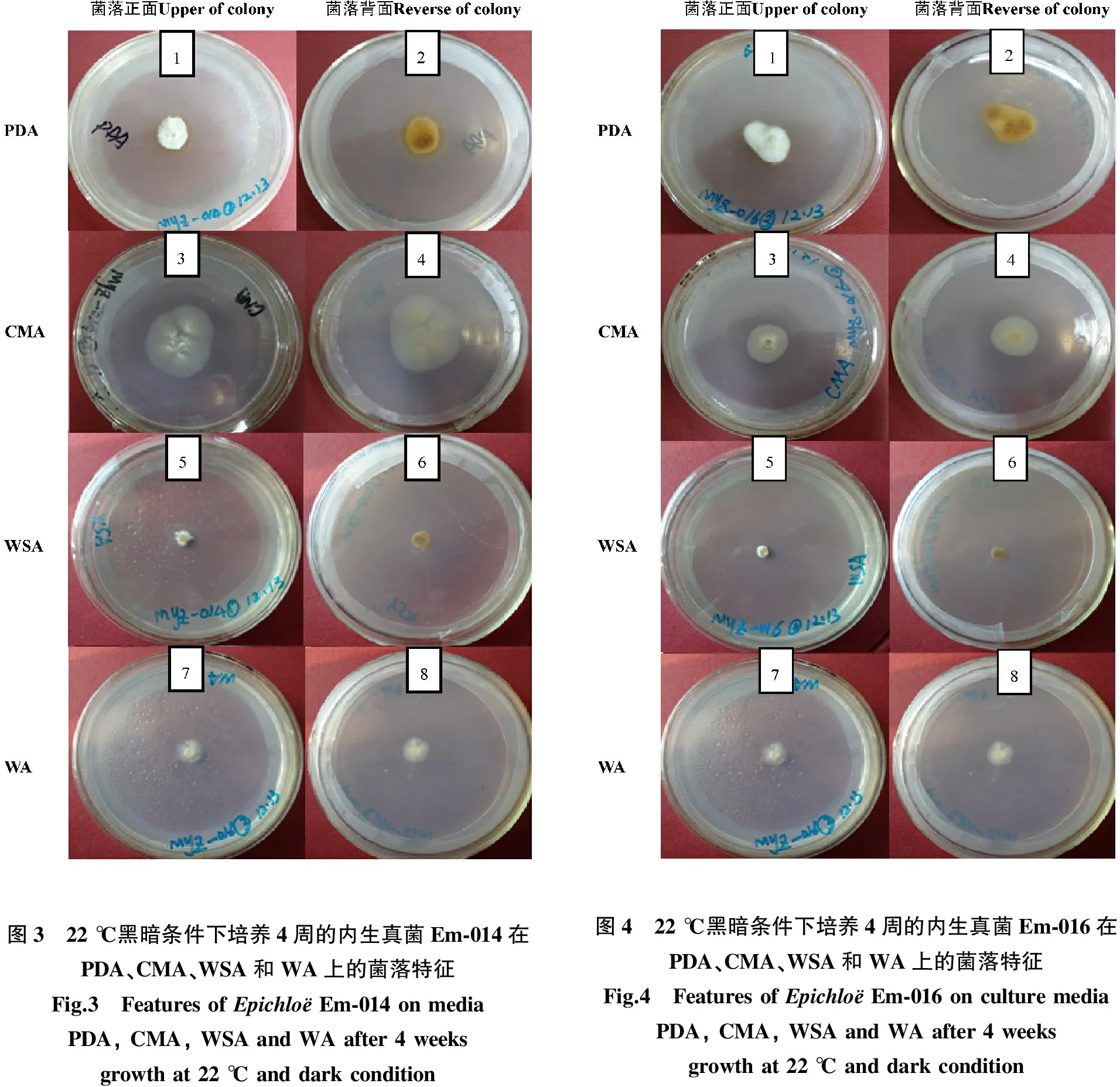
图3 22℃黑暗条件下培养4周的内生真菌Em⁃014在PDA、CMA、WSA和WA上的菌落特征Fig.3 FeaturesofEpichloёEm⁃014onmediaPDA,CMA,WSAandWAafter4weeksgrowthat22℃anddarkcondition图4 22℃黑暗条件下培养4周的内生真菌Em⁃016在PDA、CMA、WSA和WA上的菌落特征Fig.4 FeaturesofEpichloёEm⁃016onculturemediaPDA,CMA,WSAandWAafter4weeksgrowthat22℃anddarkcondition
同一培养基上不同菌株的孢子宽度存在差异。PDA上,菌株Ee-003孢子最宽;CMA上,菌株Ee-001孢子宽度最小;WSA上,菌株Ep-004和菌株Ee-003孢子宽度无显著差异(P>0.05)且显著大于其他3个菌株(P<0.05);WA上,菌株Ep-003的孢子宽度最大(表5)。
同一菌株在不同培养基上的孢子形状存在差异(表6),PDA上,Ep-004的孢子形状为椭圆形,Em-014的为近圆形,Em-016的为弯月形,Ee-001和Ee-003的孢子均为肾形;CMA上,Ep-004、Ee-001、Ee-003(图7)、Em-016的孢子均为近圆形,Em-014的为肾形。WSA上,Ep-004的孢子形状为椭圆形,Em-014(图1)和Ee-003为肾形,Ee-001和Em-016为近圆形;WA上,Ep-003孢子为肾形(图7),Ep-004为近圆形,Em-014和Em-016为椭圆形,Ee-001和Ee-003为肾形。
同一培养基上不同菌株孢子的大小也存在差异,PDA上,Ee-003的孢子最大,其他各菌株间无显著差异(P>0.05);Ep-004在CMA和WSA上孢子最大,WA上,Ep-003的孢子最大(表6)。
不同培养基上,各菌株的孢子大小趋势大致表现为:在PDA培养基上的孢子均较大,甘肃臭草内生真菌的菌株在WSA上的孢子小于在其他3种培养基上的孢子大小。
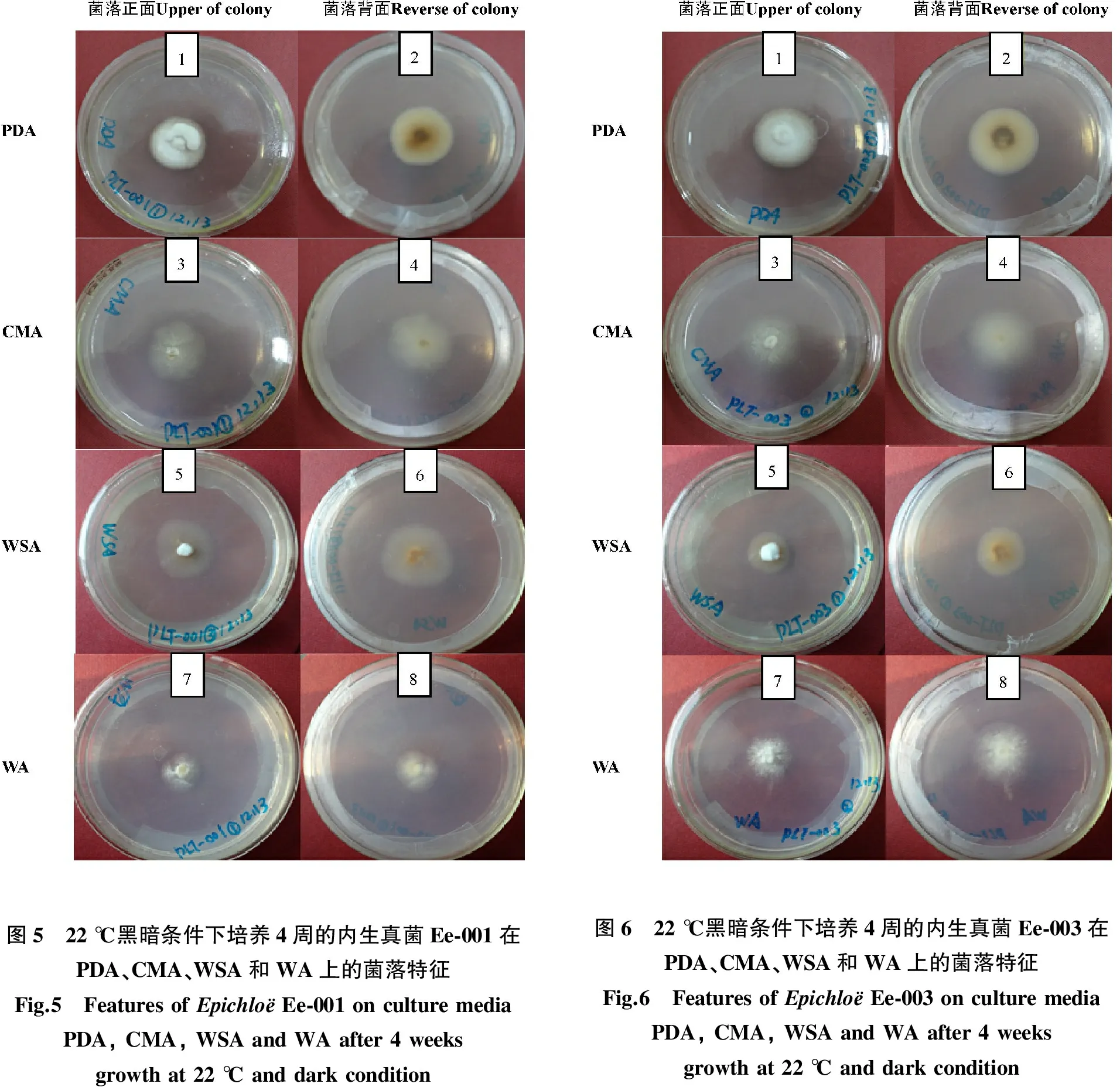
图5 22℃黑暗条件下培养4周的内生真菌Ee⁃001在PDA、CMA、WSA和WA上的菌落特征Fig.5 FeaturesofEpichloёEe⁃001onculturemediaPDA,CMA,WSAandWAafter4weeksgrowthat22℃anddarkcondition图6 22℃黑暗条件下培养4周的内生真菌Ee⁃003在PDA、CMA、WSA和WA上的菌落特征Fig.6 FeaturesofEpichloёEe⁃003onculturemediaPDA,CMA,WSAandWAafter4weeksgrowthat22℃anddarkcondition
对不同菌系的内生真菌的孢子梗长度而言,麦宾草>西藏早熟禾>甘肃臭草;对不同菌株而言,菌株Ep-004、Em-016和Ee-003在4种培养基上孢子梗的长度均不存在显著差异(P>0.05),Em-014在WSA上孢子梗最短,Ee-001在CMA上孢子梗最长,在WSA上最短(表7)。
就不同培养基来看,在PDA和WA上,5个菌株的孢子梗长度均无显著差异(P>0.05);CMA上,麦宾草内生真菌的菌株孢子梗长度无显著差异且最长,而在WSA上,这两个菌株的孢子梗长度存在显著差异(P<0.05)(表7)。
对不同菌系的内生真菌而言,孢子梗基部宽的大小为麦宾草菌系>甘肃臭草菌系>西藏早熟禾菌系;对不同菌株而言,除Ep-003外,其他5个菌株在PDA上的孢子梗基部宽分别大于其他3种培养基上的孢子梗基部宽(表8)。
从同一培养基上的不同菌株来看,在PDA上,除Em-014外,其他3个菌株的孢子梗基部宽度无显著差异(P>0.05);CMA上,Ee-001孢子梗基部宽度最大;WSA上,Ep-004的孢子梗基部宽最小;WA上Em-016的孢子梗基部宽与其他5个菌株存在显著差异(P<0.05)且最大,Ep-003、Em-014、Ee-001、Ee-003菌株的孢子梗基部宽度无显著差异(表8)。
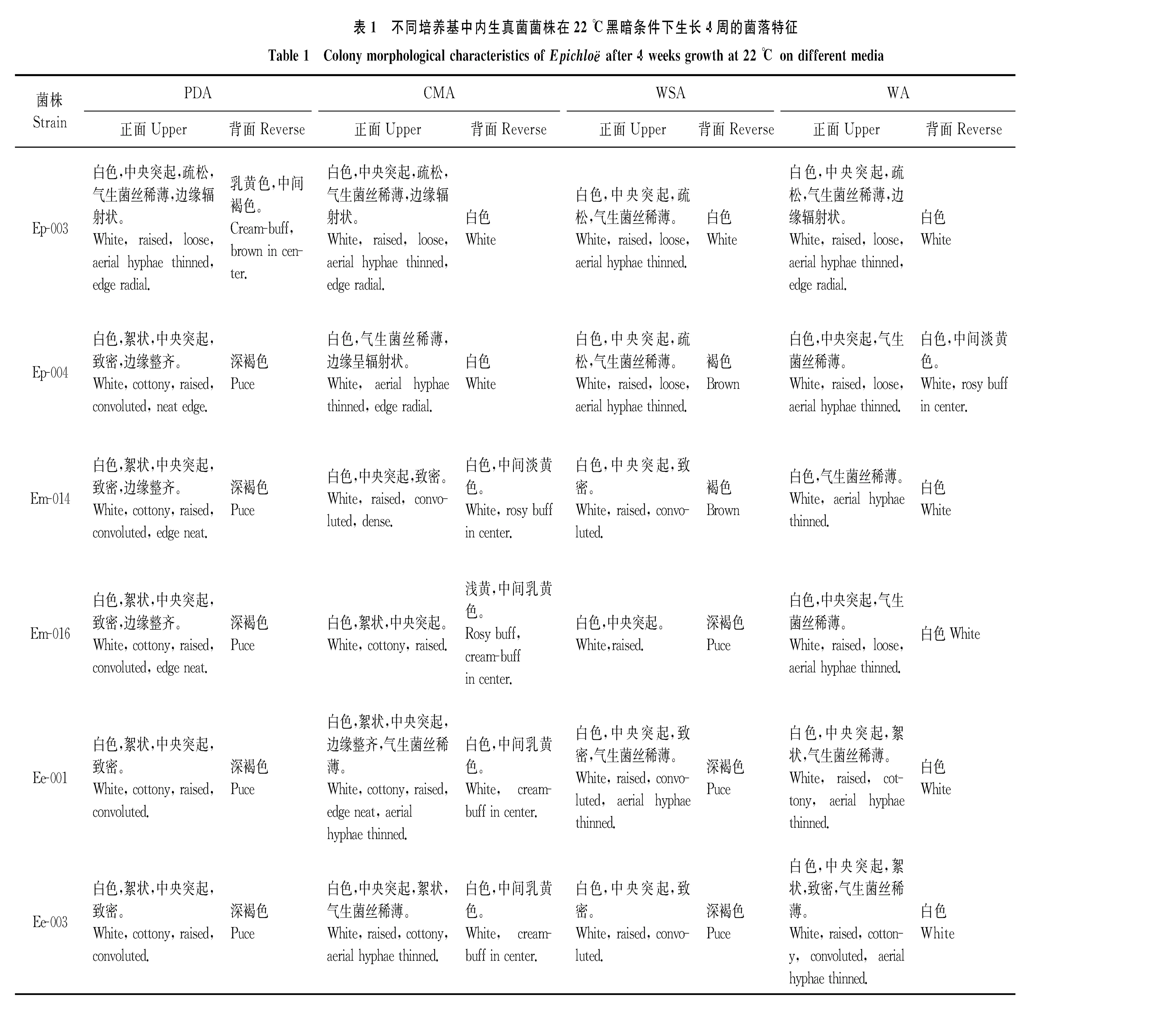
表2 不同培养基中西藏早熟禾、甘肃臭草和麦宾草内生真菌在22 ℃黑暗条件下培养4周的菌落直径
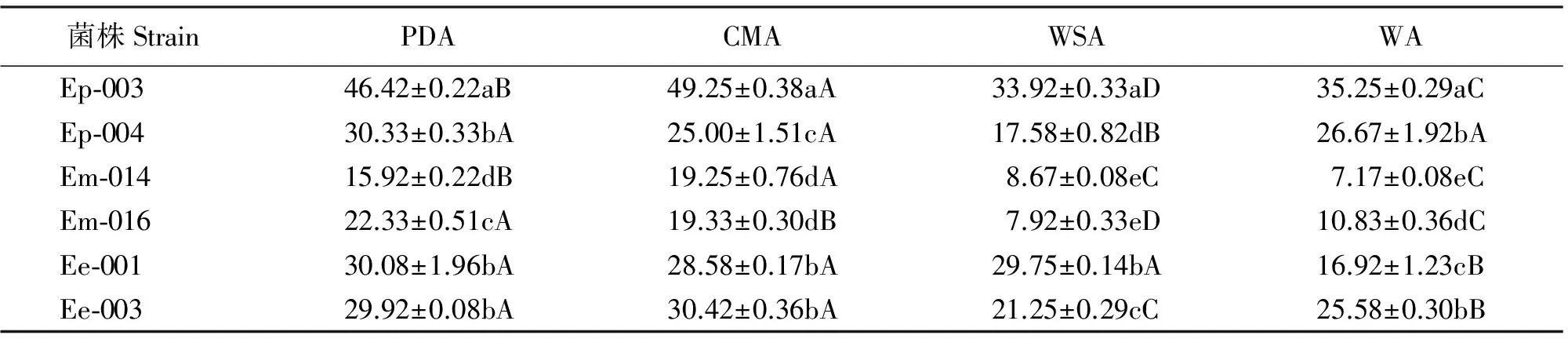
Table 2 Colony diameter of Epichloё spp isolated from Poa tibetica, Melica przewalskyi and Elymus tangutorumafter 4 weeks growth at 22 ℃ on different culture media mm
注:同行不同大写字母表示同一菌株不同培养基间差异显著(P<0.05),同列不同小写字母表示同一培养基不同菌株间差异显著(P<0.05)。表4、5、6、7、8同。
Note: Different capital letters within the same row mean significant difference among culture media at 0.05 level, and different lower case letters within the same column mean significant difference among 6 strains at 0.05 level. The same in Table 4, Table 5, Table 6, Table 7 and Table 8.

表3 不同培养基中西藏早熟禾、甘肃臭草和麦宾草内生真菌的菌株在22 ℃黑暗条件下4周后的产孢特性
注:-表示无数据居。
Note: -,no spore.
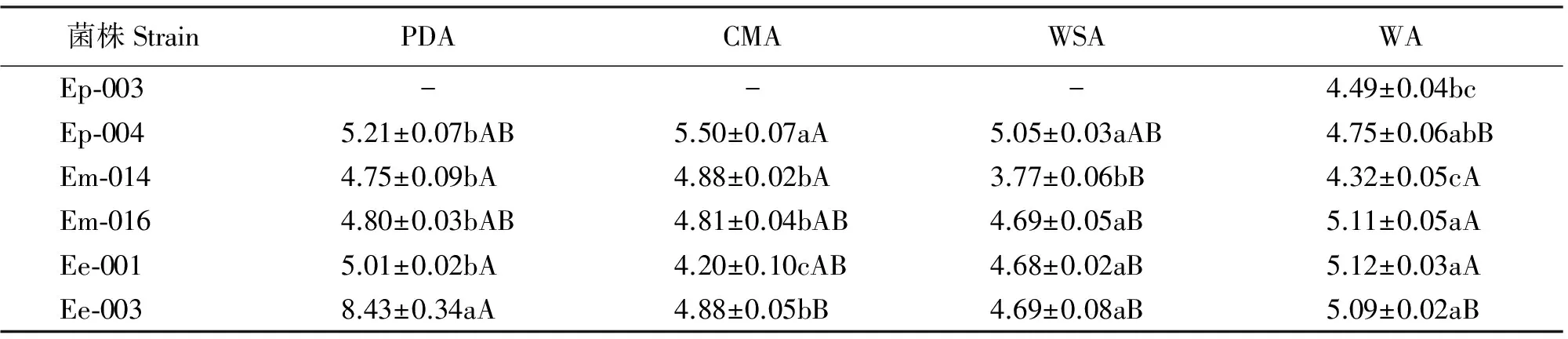
表4 不同培养基上22 ℃黑暗条件下生长4周的6个菌株孢子长度

表5 不同培养基上22 ℃黑暗条件下生长4周的6个菌株孢子宽度

图7 22 ℃黑暗条件下生长4周的内生真菌菌株的孢子及孢子梗Fig.7 Spores and sporophores of Epichloё after 4 weeks growth under 22 ℃ and dark condition 表6 不同培养基上22 ℃黑暗条件下生长4周的菌株的孢子大小及形状 Table 6 Size and shape of spores of 6 strains after 4 weeks growth under 22 ℃ and dark conditions on different culture media
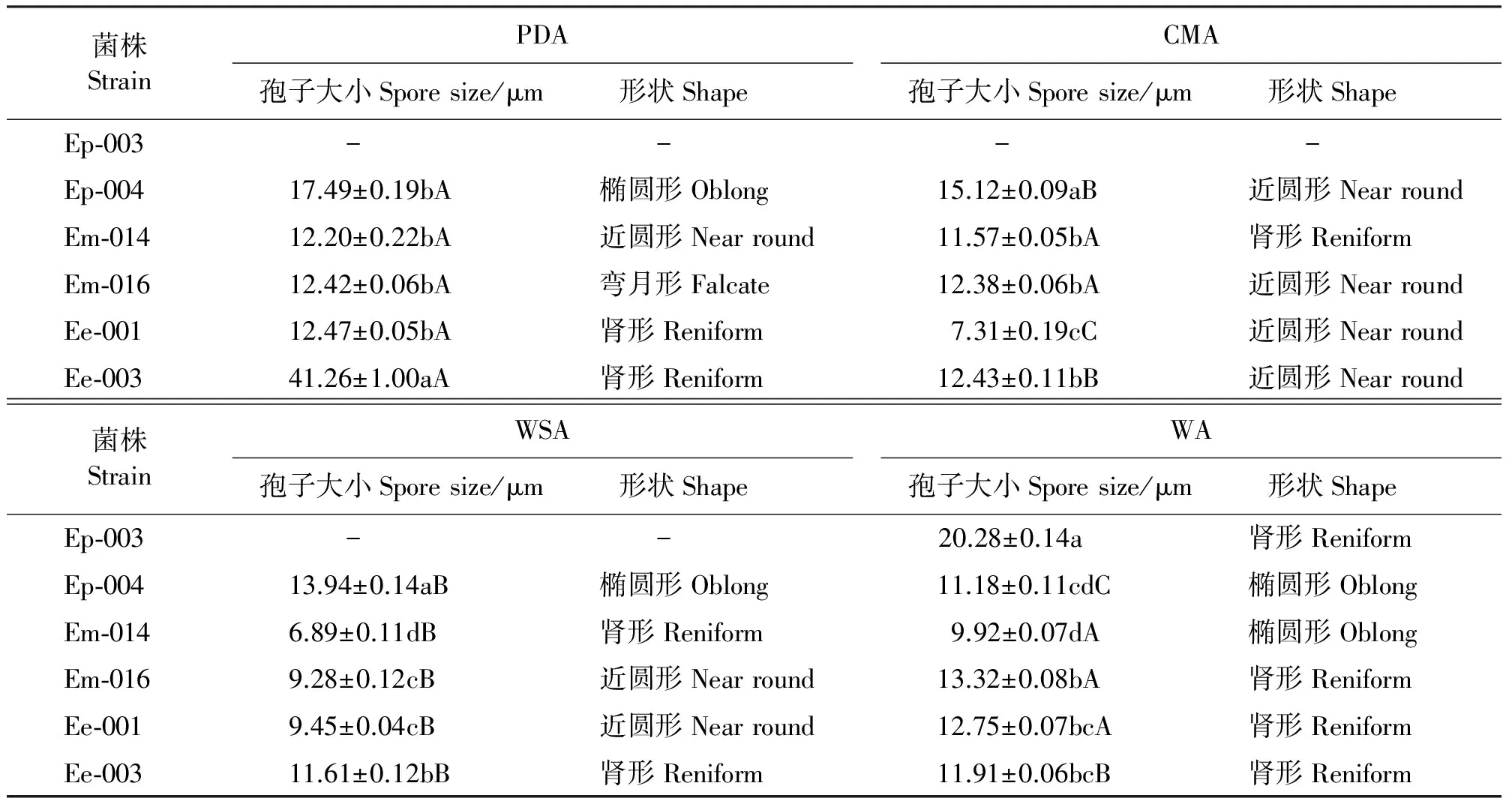
菌株StrainPDA孢子大小Sporesize/μm形状Shape CMA孢子大小Sporesize/μm形状Shape Ep⁃003 - - - -Ep⁃00417.49±0.19bA椭圆形Oblong15.12±0.09aB近圆形NearroundEm⁃01412.20±0.22bA近圆形Nearround11.57±0.05bA肾形ReniformEm⁃01612.42±0.06bA弯月形Falcate12.38±0.06bA近圆形NearroundEe⁃00112.47±0.05bA肾形Reniform7.31±0.19cC近圆形NearroundEe⁃00341.26±1.00aA肾形Reniform12.43±0.11bB近圆形Nearround菌株StrainWSA孢子大小Sporesize/μm形状Shape WA孢子大小Sporesize/μm形状Shape Ep⁃003 - -20.28±0.14a肾形ReniformEp⁃00413.94±0.14aB椭圆形Oblong11.18±0.11cdC椭圆形OblongEm⁃0146.89±0.11dB肾形Reniform9.92±0.07dA椭圆形OblongEm⁃0169.28±0.12cB近圆形Nearround13.32±0.08bA肾形ReniformEe⁃0019.45±0.04cB近圆形Nearround12.75±0.07bcA肾形ReniformEe⁃00311.61±0.12bB肾形Reniform11.91±0.06bcB肾形Reniform
3 讨论
不同培养基对禾草内生真菌的生长有一定的作用,研究发现,黑麦草内生真菌在燕麦粉琼脂培养基(OMA)上的生长优于PDA和CMA,原因是OMA培养基营养较为丰富,利于内生真菌菌落的生长[16]。本研究中6个菌株均在CMA及PDA培养基中生长优于WSA和WA培养基,原因是前两种培养基的营养较后者丰富。

表7 不同培养基上22 ℃黑暗条件下生长4周的6个菌株孢子梗长度
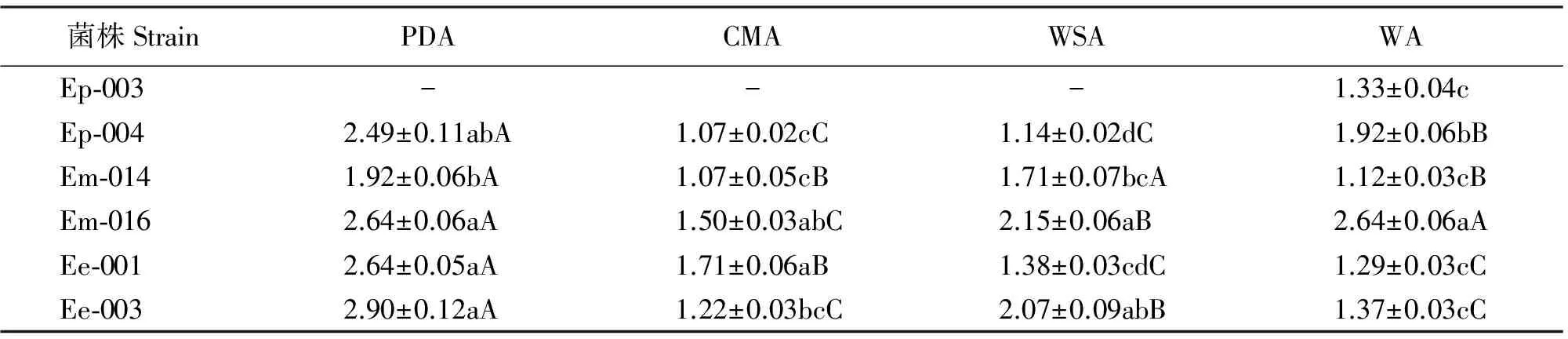
表8 不同培养基上22 ℃黑暗条件下生长4周的6个菌株孢子梗基部宽
不同培养基对禾草内生真菌的菌落形态特征产生了一定的影响,如Ep-003在PDA培养基上菌落背面为乳黄色,中间褐色,而在其他3种培养基上均为白色;Ep-004在PDA培养基上菌落正面致密,絮状,而在WSA培养基上疏松。同一培养基上,不同菌株之间也存在差别,WA培养基上,Ep-003和Ep-004菌落表面疏松,而Em-014、Em-016、Ee-001和Ee-003菌落表面均致密。PDA培养基上48株中华羊茅(Festucasinensis)内生真菌菌株的形态特征及生长特性表现出较高的多样性[17],这与本研究中6个菌株在4种培养基上的形态特性同样表现出了多样性较为类似,说明内生真菌菌株在相同条件下表现出较多的外部特征,这很可能也是因为与其具有的较为多样的外在表现有关。
高羊茅(F.arundinacea)内生真菌在WA培养基上产孢量最大,在CMA和SNA两种培养基上产孢较多,而在DMA和OMA上均产孢[16],原因是产孢属于生殖生长,而菌落生长属于营养生长,WA培养基营养贫乏,适合产孢,PDA培养基营养较好,不利于产孢。本研究中,在PDA和CMA培养基上产孢较少,而在WA培养基上产孢较多。
分离自西藏早熟禾的菌株Ep-003只在WA培养基上产孢,而Ep-004在4种培养基上都产孢,原因可能是同一菌系的不同菌株之间存在遗传多样性,但是需要更进一步的研究。
WSA培养基以前只用于病原菌的促产孢实验,但李彦忠[18]发现WSA适于埃里砖格孢病原真菌的生长与产孢。本研究首次采用WSA培养基对禾草内生真菌的产孢特性进行研究,结果表明,大多数菌株在WSA上产孢量大于WA,其原因可能是WSA中含有禾本科植物的某些特定成分,这种物质有利于促进内生真菌的产孢。
由于Epichloё属内生真菌的数量较多,所以有关不同种内生真菌在其生理特征和形态特征上存在多样性[19-20]。同时,Sun等[21]对剪股颖(Agrostismatsumurae)内生真菌在PDA上的产孢特性进行研究发现,不同菌株在同一培养基上所产生的孢子大小各不相同。本研究中各菌株的分生孢子大小,从孢子长、孢子宽、孢子梗长到孢子梗宽,在不同培养基上也存在多态性,这说明菌株的表型特征除了受到培养基的营养成分影响外,还取决定于其自身的遗传组成[22]。
[1] Siegel M R,Latch G C M,Johnson M C.Fungal endophyte of grasses[J].Annual Review of Phytopathology,1987,25:293-315.
[2] Leuchtmann A,Bacon C W,Schardl C L,White J F,Tadych M.Nomenclatural realignment ofNeotyphodiumspecies with genusEpichloё[J].Mycologia,2014,106(2):202-215.
[3] Latch G C M,Christensen M J,Samuels G J.Five endophytes ofLoliumandFestucain New Zealand[J].Mycotaxon,1984,20:535-550.
[4] White J F,Morgan-Jones G.Endophyte-host associations in forage grasses.X.cultural studies on some species ofAcremoniumsect.albo-lanosa,including a new species,A.starrii[J].Mycotaxon,1987,30:87-95.
[5] Morgan-Jones G,White J F,Piontelli E L.Endophyte-host associations in forage grasses.XIII.Acremoniumchilense,an undescribed endophyte occurring inDactylisglomeratain Chile[J].Mycotaxon,1990,39:441-454.
[6] White J F,Morrow A C,Morgan-Jones G,Chambless D A.Endophyte-host associations in forage grasses.Primary stromata formation and seed transmission inEpichloёtyphina:Developmental and regulatory aspects[J].Mycologia,1991,83(1):72-81.
[7] 李春杰.醉马草-内生真菌共生体生物学与生态学特性的研究[D].兰州:兰州大学博士论文,2005.
[8] 王静,任安芝,谢凤行,魏宇昆,高玉葆.几种诱导黑麦草内生真菌产孢的方法[J].菌物学报,2005,24(4):590-596.
[9] 金文进,李春杰,南志标.中华羊茅内生真菌Neotyphodiumsp.生物学与生理学特性的研究[J].菌物学报,2009,28(3):363-369.
[10] 赵成章,龙瑞军.石羊河上游甘肃臭草型退化草地植被恢复过程[J].山地学报,2008,26(3):286-292.
[11] 徐松鹤,尚占环,龙瑞军,齐文娟.黑土滩退化草地、高寒湿地及其交错区植物群落结构多样性[J].草原与草坪,2007(4):45-49.
[12] Li X Z,Li C J,Nan Z B.Detection and isolation of endophytes fromMelicaprzewalskyiandPoatibetica[A].Proceedings of the 8th International Symposium on Fungal Endophyte of Grasses[C].Lanzhou,China:Lanzhou University,2012:42-45.
[13] 李阳春,张宏利.祁连山东段至兰州一线早熟禾属植物的初步研究[J].国外畜牧学——草原与牧草,1997(2):24.
[14] 南志标,李春杰.禾草-内生真菌共生体在草地农业系统中的作用[J].生态学报,2004,24(3):605-616.
[15] 刘尚武.青海植物志(第4卷)[M].西宁:青海人民出版社,1999:97-103.
[16] Li C J,Nan Z B,Li F.Biological and physiological characteristics ofNeotyphodiumgansuensesymbiotic withAchnatheruminebrians[J].Microbiological Research,2008,163(4):431- 440.
[17] 杨洋,陈娜,李春杰.甘肃中华羊茅内生真菌形态多样性[J].草业科学,2011,28(2):278-283.
[18] 李彦忠.沙打旺黄矮根腐病(Embellisiaastragalisp.nov.Li&Nan)的研究[D].兰州:兰州大学博士论文,2007.
[19] Christensen M J,Latch G C M,Tapper B A.Variation within isolates ofAcremoniumendophytesfrom perennial ryegrasses[J].Mycological Research,1991,95(8):918-923.
[20] Christensen M J,Latch G M Variation among isolates ofAcremoniumendophytes(A.coenophialumand possiblyA.typhinum) from tall fescue (Festucaarundinacea)[J].Mycological Research,1991,95(9):1123-1126.
[21] Sun S.Morgan-Jones,Funk C R,White J F,Halisky P M.Endophyte-host associations in grasses.XVI.Patterns of endophyte distribution in species of the tribe agrostideae[J].American Journal of Botany,1992,79(4):472-477.
[22] 魏宇昆,高玉葆,黎川,许华,任安芝.内蒙古中东部草原羽茅内生真菌的遗传多样性[J].植物生态学报,2006,30(4):640-649.
(责任编辑 武艳培)
Effects of different medium on growth and sporulation of asexual stage ofEpichloё endophytes
LIU Li, LI Xiu-zhang, GUO Chang-hui, LI Chun-jie
(State Key Laboratory of Grassland Agro-ecosystems, College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou 730020, China)
In the present study, six fungal strains of asexual stage ofEpichloё endophytes were isolated fromPoatibetica(Ep-003, Ep-004),Melicaprzewalskyi(Em-014, Em-016) andElymustangutorum(Ee-001, Ee-003) and were cultured on different medium including potato dextrose agar (PDA), corn meal agar (CMA), wheat straw agar (WSA) and water agar (WA). The results showed that the same strain had different colonial morphologies on different medium which had the largest colonial diameter on PDA and CMA and the smallest colonial diameter on WSA and WA after four weeks growth on medium. For the performance of different strains on the same medium, the strain of Ep-003 was fastest growing with a culture diameter of 46.42 mm on PDA whereas the strain of Em-014 was slowest growing with a culture diameter of 7.07 mm on WA. There were significant difference between sporulation on different medium(P<0.05). Ep-003 only sporulated on WA and the other five isolates sporulated more on WSA and WA than on PDA and CMA. There was no significant differences between the spore length or width on different medium except with that Em-014 had the longest spore length of 3.77 μm on WSA, and Ee-003 had the longest spore length of 8.43 μm on PDA. In terms of the length of conidiophore, there was no significant differences between the length of Ep-004, Em-016 and Ee-003 on four different medium. Em-014 had the longest length of 16.74 μm on WA and the shortest length of 14.09 μm on WSA. However, Ee-001 had the longest length of 18.59 μm on CMA and the shortest length of 13.40 μm on WSA.
symbiotic fungi; conidium; growth rate; characteristics of sporulation
LI Chun-jie E-mail:chunjie@lzu.edu.cn
10.11829j.issn.1001-0629.2014-0299
2014-06-20 接受日期:2014-10-27
国家973计划课题(2014CB138702);国家自然科学基金项目(31372366)第一作者:柳莉(1987-),女,甘肃白银人,在读硕士生,主要从事禾草内生真菌共生体研究。E-mail:lliu13@lzu.edu.cn
李春杰(1968-),男,甘肃镇原人,教授,博士,主要从事禾草内生真菌共生体研究。 E-mail:chunjie@lzu.edu.cn
S432.4+4;Q939.96
A
1001-0629(2015)06-0859-11
柳莉,李秀璋,郭长辉,李春杰.不同培养基对禾草内生真菌Epichloё生长与产孢的影响[J].草业科学,2015,32(6):859-869.
LIU Li,LI Xiu-zhang,GUO Chang-hui,LI Chun-jie.Effects of different medium on growth and sporulation of asexual stage ofEpichloё endophytes[J].Pratacultural Science,2015,32(6):859-869.
