施肥处理在高温季节对闽楠幼林叶绿素荧光特性的影响
2015-05-22文仕知郭文平颜家雄杨佳伟
颜 珣 ,文仕知 ,郭文平 ,颜家雄 ,焦 帅 ,杨佳伟
(1.中南林业科技大学,湖南 长沙 410004;2.株洲市黄丰桥林场,湖南 株洲 412300;3.北京工业大学,北京 100124)
光合作用的原初反应阶段,也就是植物将吸收的光能进一步传递、转换为植物体内电能的过程中,一部分光能会损耗,并且是以荧光的方式释放出来。叶绿素荧光能够直接反映植物自身的对光能的利用和光合效率,同光合作用的关系十分密切。植物叶片的初始荧光(F0)、最大荧光(Fm)、可变荧光(Fv)、PSⅡ最大光化学量子产量(Fv/Fm)和量子产量(Yie)等多种叶绿素荧光参数可用来研究多种植物对环境胁迫的响应,同时也可根据不同种源的荧光参数进行种源与良种选择。吴建慧等通过研究两个山茶品种荧光参数日变化对春季低温的响应,得到两个山茶的抗寒性比较[1];李威通过对生长在不同光环境下东北红豆杉幼树叶片光合特性以及叶绿素荧光特性的研究,以了解东北红豆杉对光强的适应性[2]。
闽楠又称楠木,是楠木属的一个树种,属樟科,是中国特有的名贵建筑用材,国家二级保护植物[3]。至今为止,人们对闽楠这一珍贵树种的开发利用较少,研究主要集中在闽楠种群结构和空间布局[4]、种子休眠和萌发[5]、人工林生物量结构[6]、引种栽培[7]、育苗造林[8]、及光合生理特性[9]等方面的研究。
本研究以3年生闽楠幼苗为实验材料,设置4个施肥处理,一个空白样地对照,对闽楠幼林的叶绿素荧光参数进行了田间活体测定,比较不同施肥处理对其荧光特性的影响,以期阐明闽楠幼苗光合作用对不同施肥处理的响应机制,寻求高产优质的施肥用量,为苗木培育研究提供基础数据。
1 材料与方法
1.1 试验地概况
该试验地位设置于攸县黄丰桥林场,位于湖南省株洲市 , 地理位置为 113°40′E,27°18′N,为典型的亚热带季风气候。年平均气温17.8 ℃,夏季平均气温34.5 ℃,无霜期292 d,年降水量1 410 mm左右。实验样地大小均为20 m ×20 m,每个样地内均匀分布大约80株闽楠幼林,平均苗高约为60.5 cm,地径约为10.36 mm。样地坡向分别为东坡,东南破,南坡,每种处理的重复样地由西至东排开。样地内成土母岩为板页岩,土壤以板页岩发育而成的山地黄壤为主,腐殖质层厚8 cm,土壤厚度80 cm,海拔为530 m。样地具体情况如表1。
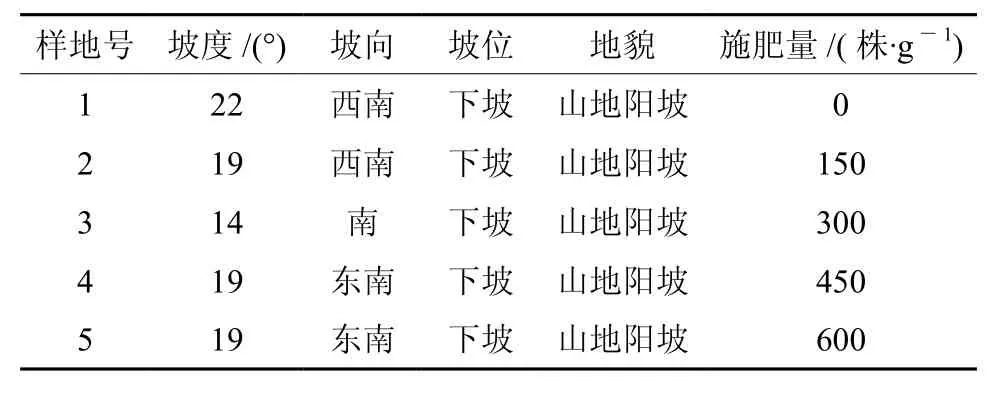
表 1 样地情况Table 1 Characteristics of sample plots
1.2 试验材料
试验材料为3年生闽楠幼林。试验采用有机复合肥,含有机质28.88%,N11.81%,有效P2O53.06%,K2O 3.37%,Pb 60.00 mg/kg,Cr 40.00 mg/kg,As 1.64 mg/kg,Hg 0.016 mg/kg。 于 2013年 7月下旬至2014年7月至9月,每月下旬于晴朗无风天气,采用德国WALZ公司MINI-PAM(超便携式调制叶绿素荧光仪)对不同施肥处理下闽楠幼林成熟功能叶片的叶绿素荧光参数进行测定。
1.3 试验方法
于上午8:00~11:00,每个种源选择3株生长健康、长势一致、光照均一的个体,随机分布于样地中。每株选择3片健康完整且充分展开的成熟未衰老叶片,测定时选择叶片中部并避开叶片的主脉。暗反应30 min后进行饱和光强处理,测定F0(初始荧光)、Fm(最大荧光)。各参数每个处理选用3个植株,每个植株选3片叶,每片叶重复测定3次,每个处理重复3次。测定F0(初始荧光)、Fm(最大荧光)和Yie(PSⅡ总的光化学量子产量)参数日变化从8:00~18:00每2 T测定一次,每个处理选用3个植株,每个植株选3片叶,每片叶重复测定3次,每个处理重复3次。
2 结果与分析
2.1 生长期F0(最小荧光)日变化
F0(初始荧光)是PSII反应中心全部打开时的荧光值即QA(PSII反应中心的电子受体)全部氧化时的荧光水平[10],PSII天线的热耗散会导致根据初始荧光降低,PSII反应中心的可逆失活或被破坏会导致初始荧光上升[11],因此可根据F0的变化来推测PSII反应中心的状况和可能的光保护机制。7月、8月、9月植物处于生长旺盛期,4种施肥处理对闽楠幼林叶片F0日变化的影响如图1所示。不同施肥处理的闽楠幼林叶片F0变化规律大致相似,均呈先降后升再趋于平衡的规律。各种施肥处理的闽楠幼林的F0值在早上8:00至中午12:00点基本呈均速下降趋势,在12:00点降至最低后又呈回升趋势,然后在下午14:00至18:00缓慢下降。7月、8月、9月夏季室外温度较高,8:00~12:00间不同施肥处理闽楠幼林F0全部呈迅速下降趋势,这种条件下的光抑制是植物自身光保护的调节机制,并非植物叶片光合组织受到强光破坏。12:00后部分样地F0有所回升,但仍低于早上8:00F0,可能是由于植物在受到高温强光胁迫后,PSII反应中心出现短暂失活。

图1 7、8、9月Fo日变化Fig.1 Dalily changes of Fo in July, August and September
2.2 Fm(最大荧光)日变化
Fm(最大荧光)是 PSII反应中心全部关闭时的荧光,反映了通过反应中心的电子传递情况,它的降低代表光抑制的一个主要特征[12],Fm大小与 QA的氧化还原状态有关[13],所以大量研究中都以Fm的变化作为判断光抑制的指标[14-15]。
不同施肥处理闽楠幼林叶片生长旺盛期Fm日变化的影响如图2所示:各施肥处理Fm日变化大致都呈先降低后升高趋势,5个处理的Fm最小值均出现于中午12:00;12点之后Fm值开始回升,14:00迅速上升到高点后后,Fm值略有下降,但是始终高于12:00的最小值,最后Fm值在18:00又上升到一个小高峰。
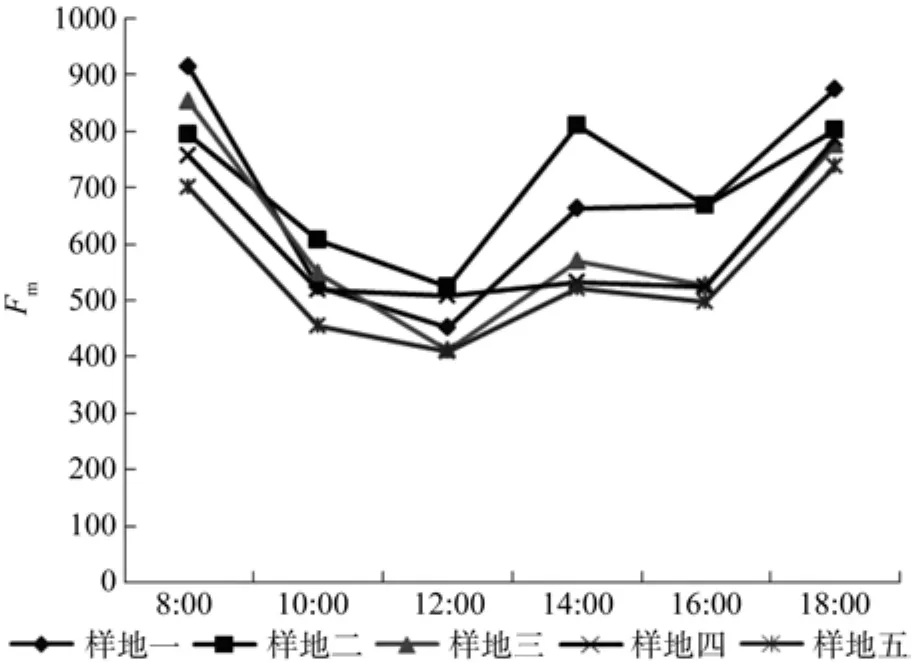
图2 7、8、9月Fm日变化Fig.2 Dalily changes of Fm in July, August and September
2.3 Yie(PSⅡ总的光化学量子产量)日变化
Yie为实际光化学量子产量,它反映PSII反应中心在部分关闭情况下的实际原初光能捕获效率,能准确地反映实际的PSII中心进行光化学反应的效率情况[11-12]。
不同施肥处理闽楠幼林叶片生长旺盛期Yie日变化的影响如图3所示:样地1、样地2、样地3、样地4、样地5都是从8:00至12:00降至一天中Yie最低点,12:00至18:00均为“升-降-升”。
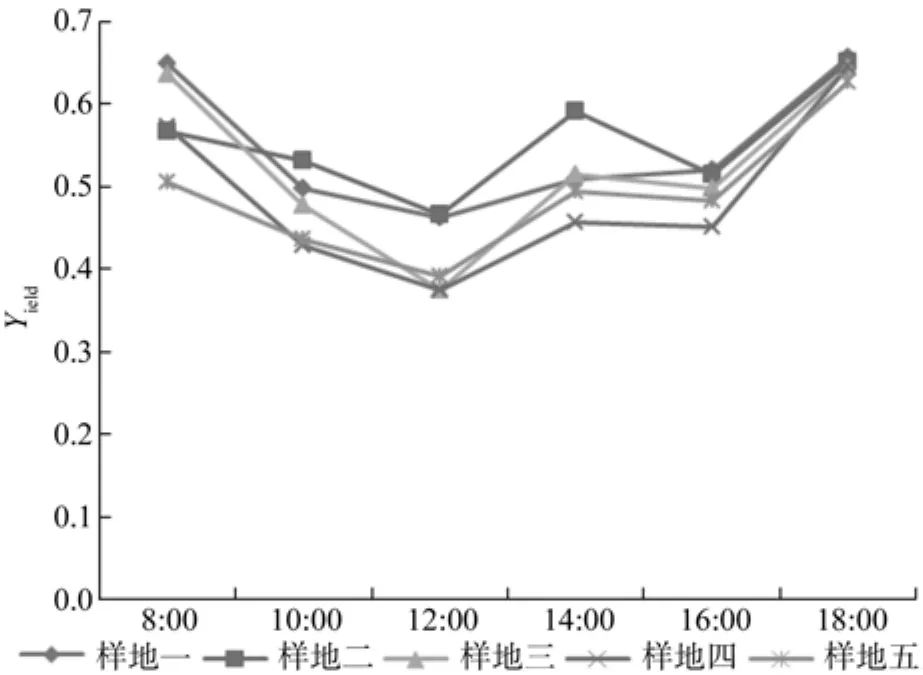
图3 7、8、9月Yie日变化Fig.3 Dalily changes of Yie in July, August and September
2.4 不同施肥处理对闽楠幼林叶片荧光参数差异比较
由表2可得,各施肥处理间叶绿素荧光参数差异较大。7月份,最大F0为639.33,最小F0为264.67,前者比后者大58.60%;最大Fm为2 495.67,最小Fm为1 310.00,前者比后者大47.51%,最大Fv/Fm为0.80,最小Fv/Fm为0.69,前者比后者大13.75%;8月份最大F0为622.67,最小F0为440.67,前者比后者大29.23%,最大Fm为2 672.00,最小Fm为2 030.33,前者比后者大24.01%,最大Fv/Fm为0.82,最小Fv/Fm为0.69,前者比后者大15.85%;9月份最大F0为618.67,最小F0为508.33,前者比后者大17.84%,最大Fm为2 444.33,最小Fm为2 200.67,前者比后者大9.60%,最大Fv/Fm为0.79,最小Fv/Fm为0.74,前者比后者大6.33%。

表 2 不同主要叶绿素荧光参数方差分析Table 2 Variance analysis of different main chlorophyll fluorescence parameters
Fm、Fv/Fm是衡量叶绿素荧光特性最常用的主要参数,因而根据可变荧光Fv在总的最大荧光Fm=Fv+F0中所占的比例Fv/Fm,即可地得出植物PSⅡ原初光能转换效率。本试验中,3号样地Fm、Fv/Fm均高于其他样地,反映了不同施肥处理间闽楠幼林PSII活性及其光化学效率的差异。
3 结论与讨论
初始荧光F0是PSII反应中心全部开放时的荧光[18]。赵丽英[19]等在研究中表明则PSII天线色素内的最初光子密度、叶绿素含量可以决定F0的大小。许大全[20]等则认为天线色素的热耗散增加是由于FQ的减少,热耗散是消耗过剩光能的重要途径,通过热耗散消耗多余光能以避免光合机构遭受光破坏。在本试验中,7月、8月、9月植物生长旺盛期不同施肥处理闽楠幼林F0中午12:00时明显低于8:00及18:00时,与上述研究结论相吻合。样地3的F0值各时期均为最低值,说明在光照温度相同的情况下,施肥处理3的施肥量最佳。
最大荧光(Fm)是PSII反应中心全部关闭时的荧光[21]。李晶[22]等研究表明,Fm与叶绿素含量呈正相关,随着叶绿素含量的进一步提高,光合作用强度增加,光化学量子产量增加,本试验中,12:00Fm降到最低,可能是由于受到高温胁迫,光合作用强度降低,植物通过热耗散保护光合机构。且本研究中可得,Fm大小于Yie呈正相关。
较高的Yie有利于提高光能转化效率,为暗反应的光合碳同化积累更多所需的能量,以促进碳同化的高效运转和有机物的积累[3]。本研究中,不同施肥处理闽楠幼林Yie的日均值都表现为样地3施肥处理相对较高,与Fm日变化一致,说明其PSII活性及其光化学效率更高,有利于光合色素把所捕获的光能以更高的速度和效率转化为化学能,从而为碳同化提供更加充足的能量,有利于光合速率的提高。
[1] 吴建慧,周蕴薇,戴思兰. 两个山茶品种光合荧光参数日变化对春季低温的响应[J]. 经济林研究,2012,30(1):40-44.
[2] 李 威,赵雨森,周志强,等. 不同光环境下东北红豆杉幼树叶光合和荧光特性比较[J]. 经济林研究,2012,30(1):51-55+94.
[3] 董春英,陈明皋,黄守成,等. 闽楠大田播种育苗及富根壮苗培育技术研究[J]. 中国农学通报,2014,16:48-52.
[4] 葛永金,王军峰,方 伟,等.闽楠地理分布格局及其气候特征研究[J].江西农业大学学报,2012,04:749-753+761.
[5] 张 伟.闽楠种子萌发技术与幼苗荧光特性的研究[D].中南林业科技大学,2013.
[6] 邵明晓,文仕知,何功秀,等. 不同林龄闽楠人工林生物量结构特征[J]. 中南林业科技大学学报,2014,34(6):44-48.
[7] 廖 柱,戴万泽,刘树根,等. 闽楠引种栽培试验研究[J]. 现代农业科技,2010,22:185+189.
[8] 黄健翔,曾火根,欧阳秋桂,等.闽楠育苗及造林技术[J].国土绿化,2013,02:43-44.
[9] 姚振一. 闽楠幼树光合特性研究[D].中南林业科技大学,2013.
[10] 张 洁,丁志强,李俊红,等. 水肥对大豆叶绿素荧光动力学参数及其产量的影响[J]. 土壤与作物,2013,2(3):122-126.
[11] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报,1999,04:444-448.
[12] 冯 涛,刘纪凤. 叶绿素荧光分析在研究植物光抑制中的应用[J]. 现代农业科技,2013,06:148+155.
[13] 徐德聪,吕芳德,潘晓杰. 叶绿素荧光分析技术在果树研究中的应用[J]. 经济林研究,2003,21(3):88-91.
[14] 徐德聪. 不同品种美国山核桃叶绿素荧光特性的研究[D].中南林学院,2004.
[15] Thomas D S, Turner D W. Banana (Musa sp.) leaf gas exchange and chlorophyll fl uorescence in response to soll drought.shading and lamina folding[J].Scientia Horticulture,2001,90(1):93- 108.
[16] 刘 昊. 模拟酸雨对12种常绿阔叶树幼苗叶绿素荧光特性的影响[D].浙江林学院,2008.
[17] 陈建明,俞晓平,程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报,2006,01:51-55.
[18] 刘 昊. 模拟酸雨对12种常绿阔叶树幼苗叶绿素荧光特性的影响[D].浙江林学院,2008.
[19] 赵丽英,邓西平,山 仑.不同水分处理下冬小麦旗叶叶绿素荧光参数的变化研究[J].中国生态农业学报,2007,15(1):63-66.
[20] 许大全,张玉忠,张荣铣,等.植物光合作的光抑制[J].植物生理学通讯,1992.28(4):237-243.
[21] Marianne K. Burke, Dudley J. Raynal Liming in fl uences growth and nutrient balances in sugarmaple (Aces saccharum) seedlings on an acidic forest soil[J].Environmental and Experimental Botany, 1998,39105-116.
[22] 李 晶,李双双,付 驰,等. 密度和施氮水平对小黑麦叶绿素荧光特性的影响[J]. 麦类作物学报,2011,01:143-148.
[23] 徐德聪. 不同品种美国山核桃叶绿素荧光特性的研究[D].中南林学院,2004.
