江西武夷山南方铁杉生长规律及更新困难的原因解释
2014-11-06祁红艳金志农杨清培袁荣斌裘利洪施建敏欧阳明
祁红艳,金志农,杨清培* ,袁荣斌,裘利洪,施建敏,欧阳明
(1.江西农业大学/江西省竹子种质资源与利用重点实验室,江西 南昌 330045;2.江西农业大学 江西鄱阳湖流域农业生态工程中心,江西 南昌 330045;3.江西武夷山国家级自然保护区管理局,江西 铅山 334500)
南方铁杉(Tsuga chinensis var.tchekiangensisa)为我国特有的珍稀裸子植物,树高可达30 m,多生于海拔600~2 100 m的中山地带,适宜的生境是雨量多、气候温和湿润和有山峰作为屏障的山坡山谷。它不仅是珍贵用材树种与优美观赏树种,也是我国第三纪残遗特有植物、濒危种,被称为植物“活化石”[1],为国家3级保护植物。由于长期开发利用,现南方铁杉林斑块状地分布在安徽南部、福建、江西、浙江、湖南、广东北部、云南等地。因此,如何合理保护并促进南方铁杉种群发展,已成为迫切需要解决的难题。
为此,不少学者分别对南方铁杉的分布格局[2-3]、群落结构[4]、种群动态[5-6]及资源与更新状况[6]进行了研究,发现该树种在群落中多处于林冠最上层,林下幼苗、幼树极少,更新较困难[7-8],然而对更新困难的原因却研究较少。影响树木自然更新的因素较多,既有树木生长速率、种子传播与萌发力等自身因素,也有林内光照、土壤养分与水分等环境条件[9],但树木本身的生长习性是影响自然更新的根本原因。因此,本文假设南方铁杉林下更新困难与其林下生长受抑制有密切关系。
为了验证以上假设,本文在江西武夷山自然保护区选取成熟南方铁杉林样地,在群落调查的基础上分别对林冠层、下木层中的南方铁杉进行树干解析,分析其树高、胸径和材积的生长过程,比较其生长速率、快速生长时间、成熟年龄等指标,以探讨以下问题:(1)本地区成熟南方铁杉林群落结构怎样,林下更新是否困难?(2)南方铁杉生长速率怎样,它是不是慢生树种?(3)与林冠层相比,下木层南方铁杉生长是不是受抑制?通过以上问题的回答,以期深入了解南方铁杉生长规律及影响因素,对揭示其濒危机制和促进自然更新具有一定理论意义。
1 材料与方法
1.1 研究区概况
江西武夷山国家级自然保护区位于江西省上饶市铅山县,地处武夷山脉北段的西北坡,地理坐标范围为 27°48'11″~28°00'35″N,117°39'30″~117°55'47″E。保护区总面积为16 007 hm2,其中核心区为4 835 hm2,缓冲区2 021 hm2,实验区9 151 hm2,保护区内保存有1 560 hm2完整的南方铁杉天然林资源,为该地区中山温性针叶林的典型代表。林下土壤为山地棕壤,平均土层厚度为20~40 cm,腐殖质层厚5 cm[10]。
1.2 研究方法
2011年8月,在江西武夷山自然保护区猪母坑,选择典型南方铁杉成熟林(24°32'15″N,114°27'34″E,海拔1 750 m),设置3块面积为20 m×30 m样方,进行群落调查。2011,2012年台风刮倒几棵南方铁杉。本文选取其中4棵(林冠层2棵,下木层2棵)作为解析木,以2 m为一区分段进行树干解析,分别在高度为 0、1.3 m、3.6 m、5.6 m、7.6 m、9.6 m……等区分段处各取一个厚约 4~5 cm 的圆盘,带回实验室,并以5 a或10 a为一龄阶,查数各圆盘的年轮数,进行树干解析[11]。
1.3 数据处理
对各南方铁杉解析木生长过程的各项测量值,分别用Logistic、Richards、Schumacher和Roялcp生长模型[11]进行树高、胸径、材积生长曲线的拟合,筛选出相关系数最大的方程为:Roялcp生长模型,即y(t)= -atbe-ct,并进一步分析生长规律。
其中,连年生长速率是通过对y(t)求导而得,即:

Roялcp生长模型呈在一个拐点,对 y(t)求二阶导数,并令其等于0,而得最大生长速率年龄,此时,最大生长速率为:

当Z(t)=θ(t)时,树木生长达到成熟数量成熟年龄,此时
文中数据统计、分析和模拟分别由Excel 2010、SPSS 16、Matlab 7.0完成。
2 结果与分析
2.1 群落基本特征
江西武夷山典型南方铁杉天然林群落层次分明,可分为4层,即林冠层(平均高22.15 m)、下木层(平均高9.29 m)、灌木层(平均高2.40 m)和草本层(表1)。林冠层物种较少,仅2种,其中南方铁杉占绝对优势,密度为 200 ind/hm2,平均高 22.67 m,最大胸径 77.0 cm,最大树高可达 30.0 m,重要值为91.54%。下木层中物种较为丰富,有植物17种,主要为多脉青冈(Cyclobalanopsis multinervis)、南方铁杉、薄叶山矾(Symplocos anomala)、多穗石栎(Lithocarpus polystachyus)等,但南方铁杉数量明显减少,仅为117 ind/hm2,其重要值也降至15.37%。灌木层植物19种,主要是毛秆箭竹(Sinarundinaria hirticaulis)、薄叶山矾(Symplocos anomala)、莽草(Illicium lanceolatum)、岩柃(Eurya saxicola)和油茶(Camellia oleifera)等,此层中没有南方铁杉幼苗、幼树。总体看来,南方铁杉成熟林种群结构呈“倒金字塔”型(图1),属衰退型种群,自然更新不良。
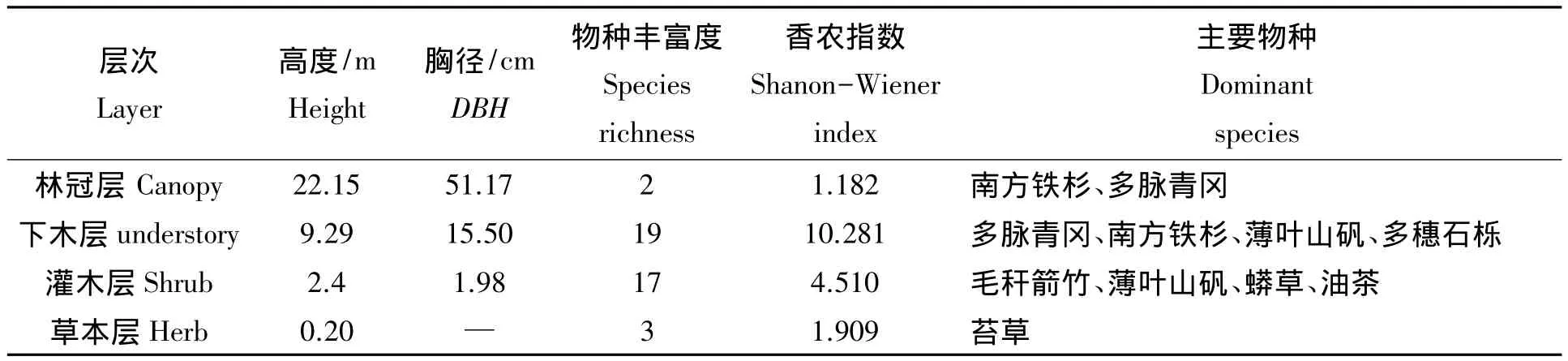
表1 南方铁杉群落的垂直结构特征Tab.1 Vertical structure of Tsuga chinensis var.tchekiangensis community
2.2 南方铁杉胸径、树高和材积总生长
根据南方铁杉解析木各龄阶的测定值,整理得其胸径、树高和材积总生长状况(图1)。结果显示林冠层南方铁杉年龄为 200 a,树高为 26.80 m,胸径(去皮)为 58.20 cm,去皮材积为 3.943 m3,带皮材积为4.125 m3。下木层南方铁杉年龄为 63 a,树高 15.10 m,胸径(去皮)为15.85 cm,去皮材积为 0.171 m3,带皮材积为 0.195 m3。
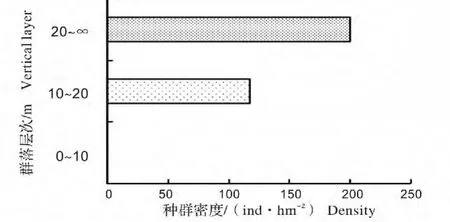
图1 南方铁杉种群结构Fig.1 Population structure of Tsuga chinensis var.tchekiangensisa in mature forest
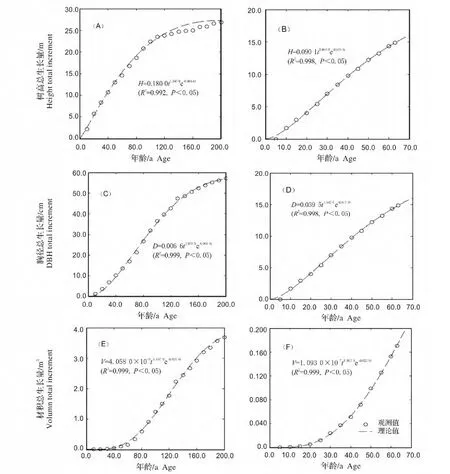
图2 南方铁杉胸径、树高和材积总生长过程Fig.2 The process of total growth of DBH,height and volume of Tsuga chinensis var.tchekiangensisa in canopy layer(A,C,E)and understory layer(B,D,F)
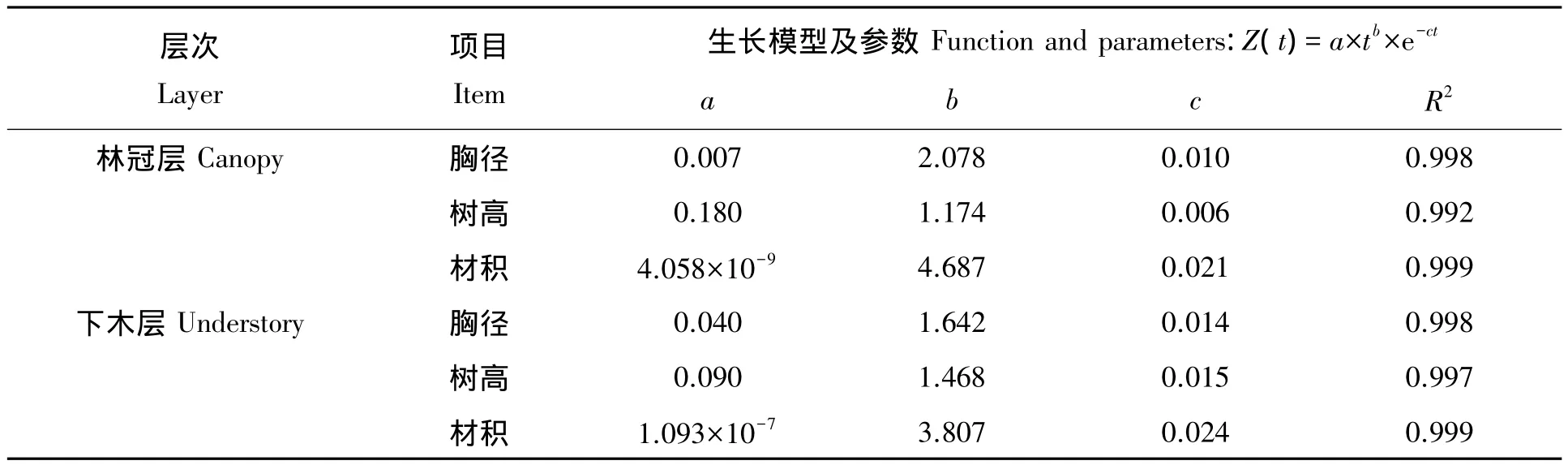
表2 南方铁杉胸径、树高和材积总生长方程及其各项参数Tab.2 The function of total increment of DBH,height and volume for Tsuga chinensis var.tchekiangensis in canopy and understory layers
再分别对林冠层和下木层的南方铁杉胸径、树高和材积总生长过程进行Roялcp生长模型拟合,求得相应参数(表2)。由表2可知,用Roялcp生长模型对林冠层和下木层的南方铁杉胸径、树高和材积总生长过程都能进行较好的效果,其相关系数R2都在0.992以上。
2.3 南方铁杉树高、胸径和材积生长过程
树干总生长过程只反映了树木不同阶段生长的总结果,要反映不同林层中不同年龄阶段的具体生长速率和生长习性,还得用连年生长速率、平均生长速率、成熟年龄等指标。
2.3.1 树高生长速率比较 林冠层和下木层南方铁杉胸径生长过程(图3)。图3可知,林冠层南方铁杉树高生长比较缓慢,前期生长速率仅0.200~0.280 m/a,第15年达到最大生长速率0.286 m/a,速生期(连年生长量≥0.20 m/a)为第1~63 a。下木层南方铁杉前期生长速率为0.13~0.31 m/a,第17年时就可达到其最大值(0.315 m/a),但速生期为第3~45 a,与林冠层相比缩短了近20 a。另外,林冠层南方铁杉成熟年龄为第29年,下木层为第30年,二者相差不大(表3)。说明不管所处群落层次与光照状况怎样,南方铁杉最大生长速率与成熟年龄出现的时间相差不大,但林下层最大生长速率较林冠层要大,且速生持续时间明显短缩。
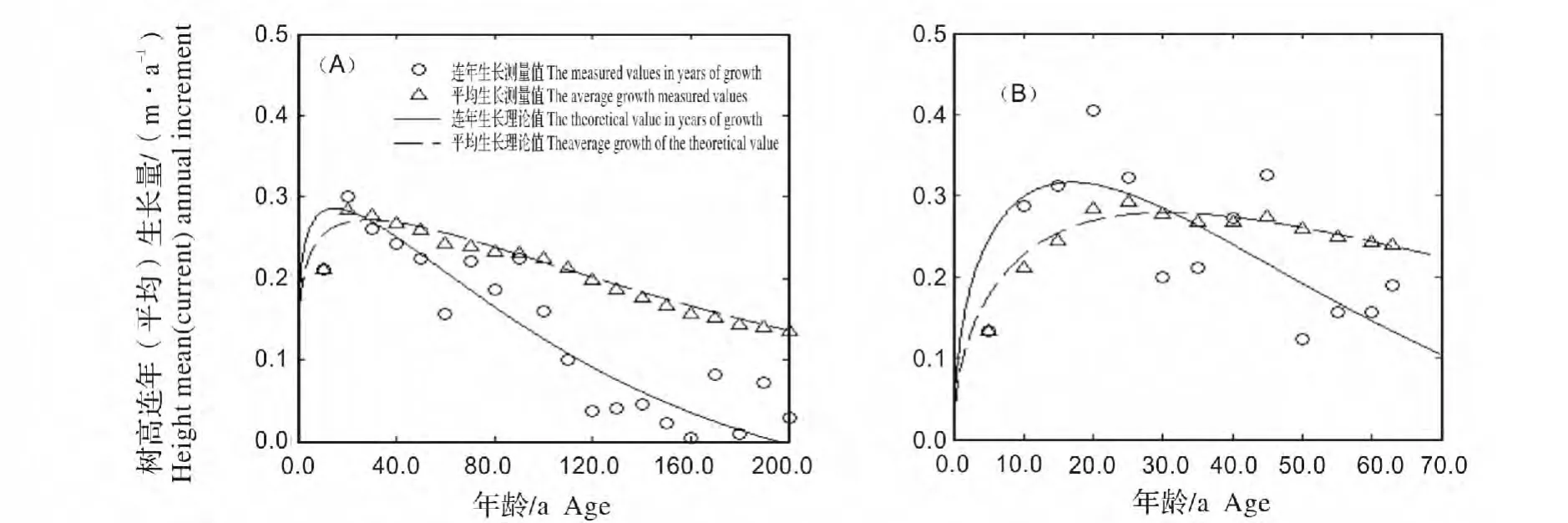
图3 林冠层与下木层南方铁杉树高生长过程Fig.3 Height growth process of Tsuga chinensis var.tchekiangensis in canopy(A)and understory(B)
2.3.2 胸径生长速率比较 林冠层和下木层南方铁杉胸径生长过程(图4)。由图4可知,林冠层南方铁杉胸径生长速度在前期生长比较缓慢,连年生长量不足0.20 cm/a,随后生长速率增加,至第65年时(对应胸径为20.40 cm)连年生长量达到最高峰为0.43 cm/a,速生生长期(按生长速率≥0.20 cm/a计)为第15~151 a。而下木层南方铁杉生长速率在第1~7 a比较缓慢,但增加相对较快,至第26年时(胸径为5.80 cm)就可达到其最大值0.29 cm/a(但比林冠层小36.83%),随后生长速率下降,其速生生长期为第8~58 a,明显比林冠层相应提前8 a,且缩短了82 a。二者连年生长量与平均生长量相交于成熟年龄,林冠层南方铁杉为第110年,而下木层为第46年,后者比前者提前了64 a(表3)。说明林下层南方铁杉胸径生长比林冠层要慢,且成熟较早,快速生长期要短。
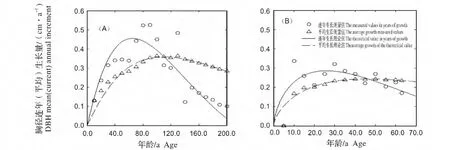
图4 林冠层与下木层南方铁杉胸径生长过程Fig.4 DBH growth process of Tsuga chinensis var.tchekiangensis in canopy(A)and understory(B)
2.3.3 材积生长速率比较 林冠层和下木层南方铁杉材积生长过程(图5)。从图5可知,林冠层南方铁杉材积生长速度在前22 a比较缓慢,连年生长量不足0.001 m3/a,此后逐渐增加,至第120年时达到最高峰为0.033 m3/a,此时对应胸径值为42.59 cm;下木层前期生长速率也较为缓慢,前19年连年生长量都小于0.001 m3/a,因本解析木年龄仅63 a,此时暂未到达最大值,但根据模型推导,其最大值时间可能在78 a,最大值仅为0.007 m3/a,比林冠层的最大生长速率小79.45%。从速生延续时间来看,林冠层速生期(连年生长速率≥0.020 m3/a)为第72~173 a,而下木层(连年生长速率≥0.005 m3/a)开始于第50年,速生期年龄提前了12 a。另外,林冠层与下木层南方铁杉材积生长的成熟年龄分别为176 a与117 a,后者比前者提前59 a(表3)。可见,下木层南方铁杉受上层林木抑制,其材积生长速率较慢,且成熟时间明显提前。
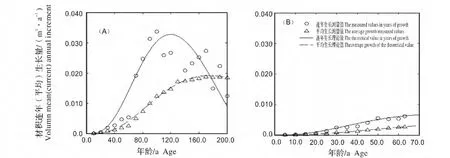
图5 林冠层与下木层南方铁杉材积生长过程Fig.5 Volume growth process of Tsuga chinensis var.tchekiangensis in canopy(A)and understory(B)
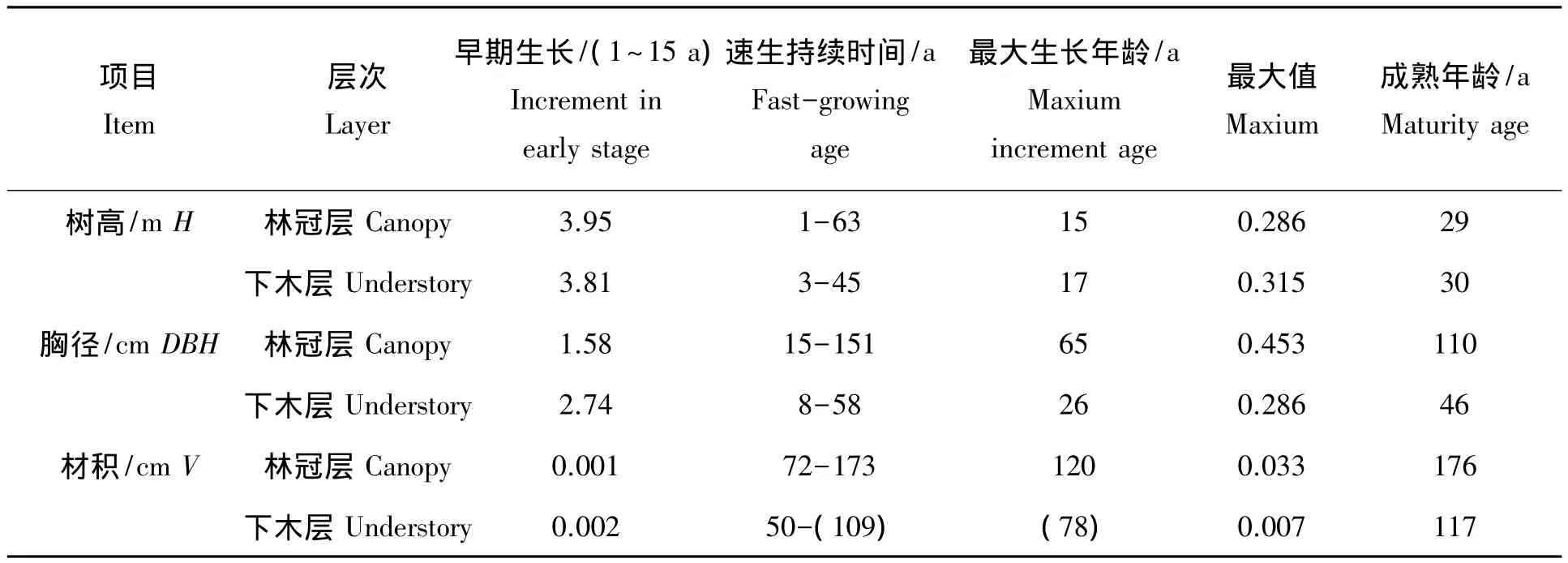
表3 林冠层与下木层南方铁杉生长速率比较Tab.3 Comparison of Height,DBH and volume increment of Tsuga chinensis var.tchekiangensis between canopy and understory
3 结论与讨论
3.1 成熟林中南方铁杉更新困难
南方铁杉为我国特有的珍稀裸子植物,是珍贵用材树种和优美观赏树种,但南方铁杉的分布面积不断收缩,生长受到抑制[6]。前人对南方铁杉种群动态与更新状况进行了广泛的研究,但结果参差不齐。黄宪刚、杜道林等认为南方铁杉自然繁殖能力已不足以维持种群的更新[7-8,12-13],郭连金[8]发现武夷山南方铁杉早在100 a前开始衰退,杜道林甚至预言600 a后南方铁杉会从贵州茂兰喀斯特山地完全消失,而张志祥[6]、袁荣斌[10]发现武夷山南方铁杉种群结构虽存在一定波动,但却能进行自然更新。本研究通过对成熟南方铁杉林群落特征分析发现,南方铁杉多位于林冠层、下木层中相对减少,而灌木层中根本没有发现幼苗与幼树,种群完全属于衰退型,说明在本研究区立地条件较为优越的地段,随着群落发育,一些阔叶树(如多脉青冈、薄叶山矾等)易于侵入南方铁杉林,林内植物增多、种间竞争加剧,南方铁杉自然更新困难。
3.2 生长缓慢是南方铁杉缺乏竞争优势的原因
对光照条件的生态适应性及生长习性决定植物竞争胜败[14]。大多数阳性植物生长速度快于阴性植物[15],但本研究发现即使是高生长较快的下木层中的南方铁杉,其最大树高生长量仅为0.453 m/a,63 a树高15.10 m,与同在武夷山区的杉木相比,南方铁杉生长缓慢得多。72 a的杉木树高可达31 m,胸径 47.5 cm,单株材积 2.4 m3,最大连年高生长达 0.98 m/a,最大胸径生长可达 1.4 cm/a,最大材积生长达0.047 m3/a[16]。同样,在广西猫儿山区273 a生南方铁杉总树高、胸径、材积分别只有9.2 m,33.5 cm,0.644 m3,其树高、胸径、材积连年生长量分别为 0.65 m/a,0.23 cm/a;福建省光泽县 46 a 生南方铁杉平均树高、胸径和单株带皮材积分别为11.8 m、22.5 cm和0.252 m3,尤其是早期生长较慢,前10 a树高总生长量仅为1.78 m[17]。说明南方铁杉虽属阳性植物,但其生长缓慢,属慢生性树种[18]。可见,阳性喜光、却生长缓慢是南方铁杉在群落种间竞争过程中处于劣势的主要原因之一。
3.3 受压“早衰”是南方铁杉更新困难的重要原因
林内光照会直接影响林下植物生存和生长[19]。本研究发现,虽然下木层中南方铁杉高生长速率比林冠层有所增大,但其速生期明显短缩,所以总高度不高,很快进入林冠层,从而获得充分光照;另外下木层南方铁杉,不但胸径、材积的最大连年生长速率比林冠层小,而年龄分别提前了39 a、42 a,数量成熟年龄分别提前了64 a和59 a,速生生长时间都明显短缩,结果林下造成南方铁杉比较“瘦弱”,如果林分郁闭度进一步增加或种间竞争更加激烈,完全会有可能导致本不多见的幼(苗)树死亡[6]。这也是成熟林下幼(苗)树较少,甚至完全不出现的原因。这些结果说明了南方铁杉生长缓慢、林下受压“早衰”、甚至死亡是南方铁杉林下完成更新的重要原因之一。
3.4 人工促进更新建议
虽然南方铁杉群落普遍存在林下更新困难的问题[7-8],然而在林窗、林缘或瘠薄生境中却发现大量幼苗[10,20],说明林下更新困难的原因不是种源不足或种子生理的问题,而南方铁杉阳性喜光、生长缓慢的生物学特性,才是它在森林群落光资源竞争中取于劣势的根本原因。目前,江西武夷山自然保护区的南方铁杉一般都与阔叶树组成了多层混交林,且多处林冠层,只有少数仍居更新层或演替层。如果不加以人工辅助,林下南方铁杉生长就会受到影响,导致南方铁杉难以在成熟林分中自然更新。因此,从珍稀植物保护以及维持武夷山中山温性针叶林景观生态稳定的角度出发,建议对现有部分南方铁杉林实行科学管理[21],调整其群落结构和种群分布格局,对天然林进行适度的人为干扰(择伐、间伐),以降低林分的郁闭度,改善群落的通风条件、促进种子传播与萌发。同时,重点保护现有母树、幼树和幼苗,减少种内或种间竞争,促进幼苗、幼树生长,实现南方铁杉这一珍稀植物正常更新。
[1]李林,魏识广,黄忠良,等.猫儿山两种孑遗植物的更新状况和空间分布格局分析[J].植物生态学报,2012,36(2):144-150.
[2]封磊,洪伟,吴承祯,等.南方铁杉种群结构动态与空间分布格局[J].福建林学院学报,2008,28(2):110-114.
[3]何建源,卞羽,吴焰玉,等.不同坡向濒危植物南方铁杉的分布格局[J].中国农学通报,2010,26(13):122-125.
[4]张志祥,刘鹏,刘春生,等.浙江九龙山南方铁杉(Tsuga tchekiangensis)群落结构及优势种群更新类型[J].生态学报,2008(9):4547-4558.
[5]封磊,洪伟,吴承祯,等.珍稀濒危植物南方铁杉种群动态研究[J].武汉植物学研究,2003,21(5):401-405.
[6]张志祥,刘鹏,蔡妙珍,等.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1146-1156.
[7]杜道林,刘玉成,刘川华.茂兰喀斯特山地南方铁杉种群结构和动态初探[J].西南师范大学学报:自然科学版,1994,19(2):169-174.
[8]郭连金,洪森荣,夏华炎.武夷山自然保护区濒危植物南方铁杉种群数量动态分析[J].上饶师范学院学报:自然科学版,2006,26(6):74-78.
[9]王传华,李俊清,陈芳清,等.鄂东南低丘地区枫香林下枫香幼苗更新限制因子[J].植物生态学报,2011,35(2):187-194.
[10]袁荣斌,邹思成,兰文军,等.江西武夷山国家级自然保护区南方铁杉资源调查初报[J].江西林业科技,2012(4):37-39.
[11]孟宪宇.测树学[M].北京:中国林业出版社,2006.
[12]黄宪刚,谢强.猫儿山南方铁杉种群结构和动态的初步研究[J].广西师范大学学报:自然科学版,2000(2):86-90.
[13]冯祥麟,胡刚,刘正华.贵阳高坡南方铁杉群落特征及种群动态调查研究[J].贵州林业科技,2011(2):26-29.
[14]Lauri J S,Steven R R.Mechanisms of competition between Douglas-fir and red alder seedlings[J].Ecology,1992,73(1):30-45.
[15]Brian J P,Kurt S P.The vertical development of early successional forests in northern Michigan,USA[J].Journal of Ecology,1993,81(2):271-285.
[16]李晓铁.猫儿山林区南方铁杉生长调查初报[J].广西林业科技,1992,21(1):24-26.
[17]罗金旺.福建光泽南方铁杉天然林的生长规律与生物量[J].福建林学院学报,2011(2):156-160.
[18]俞新妥,何智英,叶再春,等.武夷山保护区天然杉木混交林考察初报[J].福建林学院学报,1983,3(1):1-25.
[19]Sven W,Holger F,Franka H.Canopy effects on vegetation caused by harvesting and regeneration treatments[J].European Journal of Forest Research,2011,130(1):17-40.
[20]钱晓鸣,黄耀坚,张艳辉,等.武夷山自然保护区南方铁杉外生菌根生物多样性[J].福建农林大学学报:自然科学版,2007,36(2):180-185.
[21]Robert T F,Klaus J P.Patterns in spatial extent of gap influence on understory plant communities[J].Forest Ecology and Management,2008,255(7):2801-2810.
