班氏跳小蜂对扶桑绵粉蚧的寄生效能
2014-06-27王竹红温玉琪福建农林大学植物保护学院农业部闽台作物有害生物综合治理重点实验室福建福州350002
高 原, 黄 建, 王竹红, 温玉琪, 钟 顺(福建农林大学植物保护学院, 农业部闽台作物有害生物综合治理重点实验室, 福建 福州 350002)
扶桑绵粉蚧PhenacoccussolenopsisTinsley(半翅目Hemiptera: 粉蚧科Pseudococcidae)是一种严重威胁大田作物、园林观赏植物、果树和蔬菜等经济作物安全生产的入侵性害虫(朱艺勇等,2011), 原产北美洲,后逐步扩展至南美洲、非洲、亚洲、欧洲和澳洲等世界各地(朱艺勇等,2011;Akintola & Ande,2008;Hodgson et al.,2008;Abbasetal.,2012)。2009年,国家农业部、国家质检总局联合发布的第1147号公告中将该虫列入中华人民共和国进境植物检疫性有害生物名录第436种检疫性有害生物(武三安和张润志,2009;马骏等,2009)。扶桑绵粉蚧寄主范围广,可为害葫芦科、豆科、茄科、唇形科、锦葵科、藜科等18科55种植物(Hodgsonetal.,2008;吴定发等,2011),对农林业生产造成重大损失。扶桑绵粉蚧主要危害植物的幼嫩部位,以雌成虫和若虫吸食植物汁液,影响植株生长,进而影响果实的产量和质量。此外,该虫还可分泌蜜露污染叶片,导致煤烟病发生,降低了叶片光合作用功能,导致受害植株生长缓慢或停止,严重时造成植株成片死亡(武三安和张润志,2009)。
自然界中,扶桑绵粉蚧的天敌较多,有捕食性天敌、寄生性天敌和致病性微生物等(吴定发等,2011;Tanwaretal.,2011;陈华燕等,2011;Hayat,2009),其中班氏跳小蜂AenasiusbambawaleiHayat(膜翅目Hymenoptera: 跳小蜂科Encyrtidae)是其优势种天敌,在扶桑绵粉蚧种群的自然控制中起着重要作用,对该害虫的生物防治具有潜在应用价值(陈华燕等,2011;Hayat,2009;Dhawanetal.,2011;Fandetal.,2011)。国内有关该寄生蜂的研究报道较少,主要集中在寄生选择、发育历期、功能反应等(黄玲等,2011;何嫏芬等,2012;Huangetal.,2012)。本文研究了在不同蜂蚧比条件下,班氏跳小蜂对扶桑绵粉蚧寄生效能的影响,并组建自然种群生命表,采用种群趋势指数(I)、干扰作用控制指数(IIPC)和排除作用控制指数(EIPC)评价班氏跳小蜂对扶桑绵粉蚧的控制作用,为扶桑绵粉蚧的生物防治提供参考依据。
1 材料与方法
1.1 试验材料
1.1.1 供试虫源 扶桑绵粉蚧和班氏跳小蜂均是福建农林大学经济昆虫种质资源保存基地饲养的种群。室内采用马铃薯苗繁殖扶桑绵粉蚧,其上接种班氏跳小蜂。
培育马铃薯苗至一定高度,用毛笔将扶桑绵粉蚧初孵若虫轻轻扫至马铃薯苗上,在温度25±1℃、RH60%-70%、光照周期12L:12D的养虫室内繁殖;待粉蚧发育至合适龄期,接种班氏跳小蜂,进行大量继代饲养繁殖。
1.2 试验方法
1.2.1 不同蜂蚧比对班氏跳小蜂寄生效能的影响 取扶桑绵粉蚧3龄雌若虫,设置5个不同密度10头、20头、30头、40头、50头/叶(马铃薯)。在保鲜盒(长:宽:高=18 cm:8 cm:5 cm)盒盖开一长方形通气口(长:宽=7 cm:3 cm),用200筛目的白纱网封住。把有不同密度扶桑绵粉蚧的马铃薯叶片放入保鲜盒中,叶柄用湿棉花外裹锡箔纸包住以保湿。
取羽化后已交配但未产卵的班氏跳小蜂雌蜂,设置3个不同密度1头、2头、3头/盒,按照不同蜂蚧比要求将蜂放入各个保鲜盒中,做好标记。试验条件为温度25±1℃、RH60%-70%、光照周期12L:12D。重复6次。待班氏跳小蜂发育为蛹期时,收集各个处理的蛹于培养皿(Ф9 cm)内,待蛹羽化为成蜂后,统计各处理的粉蚧被寄生量和出蜂量。
1.2.2 班氏跳小蜂对扶桑绵粉蚧的控制效能 组建自然种群生命表评价班氏跳小蜂对扶桑绵粉蚧的控制效能。
试验地点为福建农林大学经济昆虫种质资源保存基地田间网室,网室内种植马铃薯。
2013年9月到12月,种植马铃薯(苗高约20 cm)苗若干,设置为对照区和放蜂区。
对照区:在各个独立网室内培育马铃薯苗若干,每苗上接种扶桑绵粉蚧初孵若虫100头,未放蜂。
放蜂区:接种扶桑绵粉蚧22 d后,在网室内释放羽化当天且已经交配未产卵的班氏跳小蜂雌蜂,设置为20头、40头、60头、100头/网室4个不同处理。
每7 d调查1次,统计记录扶桑绵粉蚧(1龄、2龄、3龄若虫、伪蛹和雌成虫)的数量和死亡原因,至3个世代结束为止。
1.3 数据处理
1.3.1 不同蜂蚧比对扶桑绵粉蚧的寄生效能 随着班氏跳小蜂幼虫的发育,被寄生的扶桑绵粉蚧体表的蜡粉状物质开始逐渐脱落,最后形成僵蚧。在班氏跳小蜂羽化前期,透过褐色的僵蚧外壳可看到黑色成熟的小蜂蛹体。班氏跳小蜂羽化时会在僵蚧体上形成一个圆形的羽化孔。
寄生蜂的寄生率和羽化率计算如下:
寄生率= (羽化寄生蜂数+未羽化死在寄主体内的寄生蜂数)/供寄生的寄主数×100%
羽化率=羽化寄生蜂数/寄生蜂黑色蛹体数×100%
数据统计采用PASW Statistics 17.0软件进行处理分析。
1.3.2 扶桑绵粉蚧各虫态的作用因子存活率估算 粉蚧虫体残缺不全或变硬变干,是由捕食性天敌捕食或刺吸作用造成;虫体发黑,但身体变软,则是被菌类感染所致,与被捕食的一起归为“捕食及其它”。而扶桑绵粉蚧的2龄若虫、3龄若虫及成虫被寄生蜂寄生后会变成僵蚧,有羽化孔的则表示寄生蜂已羽化岀蜂,一起归为“寄生蜂寄生”。
按照作用因子的先后关系,1龄若虫的主要作用因子是“捕食及其它”;2龄、3龄若虫和雌成虫的主要作用因子均为“寄生蜂寄生”、“捕食及其它”。
1.3.3 扶桑绵粉蚧自然种群生命表的组建 根据庞雄飞和梁广文(1995)、庞雄飞等(1986、1988)的方法,组建以作用因子组配的自然种群生命表,建立种群趋势指数模型,分析扶桑绵粉蚧的种群发展趋势:
I = N1/N0= S1S2S3…SKFPFP♀∑Pfi(SAa) ′
其中 I:种群趋势指数; N1、N0:下一代、当代的种群数量; Si:各个作用因子相应的存活率(1,2,3…,K); F:设定的标准卵量; PF:达标准卵量的概率; P♀:雌性概率; Pfi:成虫逐日产卵概率; SAa:成虫逐日存活率。
各龄期的数量、期中值及存活率的估计参照庞雄飞和梁广文(1995)的方法。
1.3.4 干扰作用控制指数(IIPC)的计算 庞雄飞和梁广文(1995)在种群趋势指数的方程的基础上,提出了种群干扰作用控制指数(Interference of Index of Population Control,IIPC),研究各类因子对种群趋势指数的影响。干扰作用控制指数(IIPC)为:
IIPC (Si) = I′/I = S1S2S3…Si′…SKFPFP♀∑Pfi(SAa)′/S1S2S3…Si…SKFPFP♀∑Pfi(SAa)′
式中Si:各个作用因子相对应的存活率(i=l,2,3…K); F:设定的标准卵量; PF:达标准卵量的概率; P♀:雌性概率; Pfi:成虫逐日产卵概率; SAa:成虫逐日存活率。
1.3.5 排除作用控制指数(EIPC)的计算 在各个作用因子中,如果其中一个因子E被排除,该因子的相对应的存活率SE=1,种群趋势指数将改变为IE,其排除作用控制指数(Exclusive Index of Population Control,EIPC)为:
EIPC = IE/I,= l/SEIE= EIPC×I
EIPC的含义是:如排除某个因子的控制作用,种群趋势指数将增为原来种群趋势指数的EIPC倍。因此,如果某因子的排除作用控制指数(EIPC)越大,则说明该因子的控制作用越强。EIPC可以作为重要因子(Important factors)和关键因子(Key factors)的分析指标(庞雄飞等,1984;庞雄飞和梁广文,1995;庞雄飞,2000)。
如果多个因子被排除,则其总的排除作用控制指数将为被排除的因子相对应排除作用控制指数的乘积。
2 结果与分析
2.1 不同蜂蚧比条件下班氏跳小蜂对扶桑绵粉蚧的寄生率
试验中共设置1:10、 1:20、 1:30、 1:40、 1:50、 2:10、 2:20、 2:30、 2:40、 2:50、 3:10、 3:20、 3:30、 3:40、 3:50的15个蜂蚧比组合。不同蜂蚧比条件下班氏跳小蜂对扶桑绵粉蚧的寄生率如图1。

图1 不同蜂蚧比条件下班氏跳小蜂对扶桑绵粉蚧的寄生率Figure 1 The parasitic rate under different density ratio of Aenasius bambawalei to Phenacoccus solenopsis
由图1可以看出,引入1头班氏跳小蜂到5个不同密度处理的扶桑绵粉蚧3龄若虫中,当蜂蚧比为1:10时,寄生率最高,为71%;蜂蚧比为1:20时次之,为69.2%;蜂蚧比为1:50时,寄生率最低,为41%,且经Duncan检验,在0.05水平上,差异显著,同时也是所有处理中寄生率最低的。引入2头班氏跳小蜂到5个不同密度处理的扶桑绵粉蚧3龄若虫中,当蜂蚧比为2:50时,寄生率最高,达92%,经Duncan检验,在0.05水平上,差异显著,同时也是所有处理中寄生率最高的;当蜂蚧比为2: 30时,寄生率最低,为64.6%。引入3头班氏跳小蜂到5个不同密度处理的扶桑绵粉蚧3龄若虫中,蜂蚧比为3:50时,寄生率最高,为73%;蜂蚧比为3:10和3:40时,寄生率最低,为58%。
2.2 班氏跳小蜂对扶桑绵粉蚧的控制作用
组建扶桑绵粉蚧在放蜂区和对照区不同世代的自然种群生命表,见表1、表2。
在对照区,扶桑绵粉蚧第2、3代种群趋势指数分别为44.0240和43.4000;在放蜂区,扶桑绵粉蚧第2、3代种群趋势指数与对照区相比有明显降低,分别释放20头、40头、60头和100头寄生蜂后,其种群趋势指数分别为16.6403和13.7783,16.5633和11.9783,10.0356和6.0356,11.2521和7.4521;尽管种群趋势指数都大于1,但与对照区相比,种群数量呈明显下降趋势。从各个作用因子来看,第2、3代各虫期“寄生蜂寄生”因子作用下的存活率明显低于“捕食及其它”,这对降低扶桑绵粉蚧种群趋势指数起到关键作用。
比较放蜂区和对照区的IIPC,释放20头、40头、60头和100头寄生蜂后,放蜂区扶桑绵粉蚧第2代自然种群增长的数量分别下降到对照区的34.79%、29.34%、24.31%和24.52%;第3代自然种群增长的数量分别下降到对照区的30.67%、27.44%、12.82%和21.21%。可见,释放班氏跳小蜂后对扶桑绵粉蚧控制效果明显。
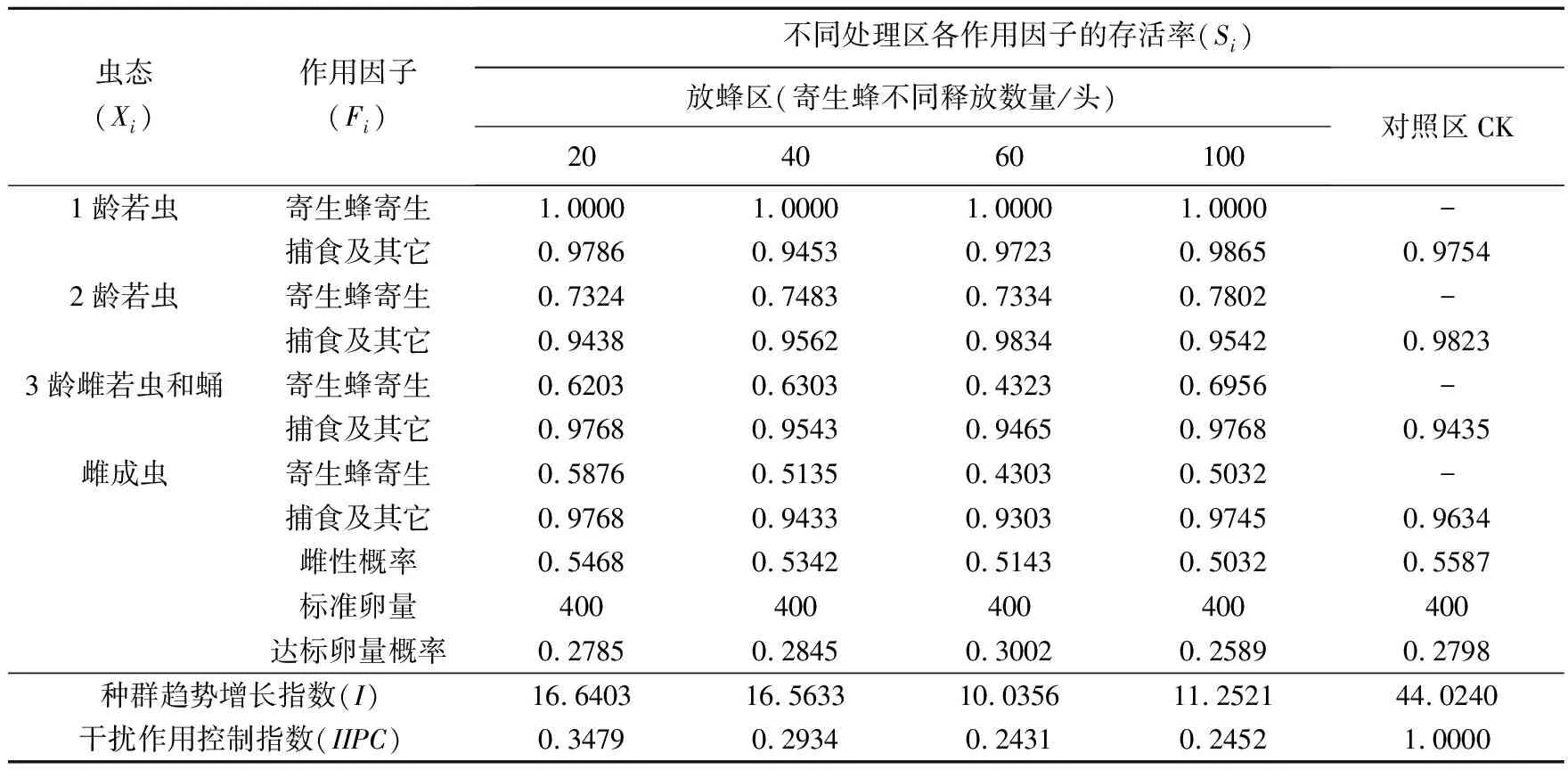
表1 扶桑绵粉蚧第2代自然种群生命表(2013年)Table 1 The life table of the 2nd generation population of Phenacoccus solenopsis(2013)

表2 扶桑绵粉蚧第3代自然种群生命表(2013年)Table 2 The life table of the 3rd generation population of Phenacoccus solenopsis(2013)
采用EIPC分析扶桑绵粉蚧第2、3代自然种群系统中的重要因子(表3、4),结果表明,“寄生蜂寄生”因子对扶桑绵粉蚧的种群数量发展起着非常重要的作用。在不同放蜂区释放20头、40头、60头和100头班氏跳小蜂后,扶桑绵粉蚧第2、3代的“寄生蜂寄生”因子的干扰指数分别为2.4213和4.0225,2.5706和3.9568,3.0629和8.5925,2.8766和5.1633,均大于“捕食及其它”因子的干扰指数,说明“寄生蜂寄生”因子是重要因子,如排除这个因子的作用,不同放蜂区(分别释放20头、40头、60头和100头班氏跳小蜂)扶桑绵粉蚧的第2代的种群趋势增长指数将会比原来分别增长至少2.4213倍、2.5706倍、3.0269倍和2.8766倍;第3代的种群趋势增长指数将会比原来分别增长至少4.0225倍、3.9568倍、8.5925倍和5.1633倍。

表3 扶桑绵粉蚧第2代各作用因子的干扰作用控制指数(2013年)Table 3 The EIPC of the 2nd generation population of Phenacoccus solenopsis(2013)

表4 扶桑绵粉蚧第3代各作用因子的干扰作用控制指数(2013年)Table 4 The EIPC of the 3rd generation population of Phenacoccus solenopsis(2013)
经过对干扰作用控制指数(IIPC)和排除作用控制指数(EIPC)的分析,可以看出释放班氏跳小蜂后,对于扶桑绵粉蚧种群的增长起着明显的控制作用。由此可见,班氏跳小蜂是应用潜力很大的优势种天敌。
3 小结与讨论
班氏跳小蜂是扶桑绵粉蚧的内寄生蜂,在印度、巴基斯坦和中国都是扶桑绵粉蚧的优势种寄生蜂(陈华燕等,2011;Pala & Saini,2011;Arifetal.,2012),对扶桑绵粉蚧种群起着重要的自然控制作用,对该害虫的生物防治具有潜在应用价值。本文研究了在不同蜂蚧比条件下,班氏跳小蜂对扶桑绵粉蚧的寄生效能,并利用自然种群生命表评价了班氏跳小蜂对扶桑绵粉的控制作用。初步筛选了班氏跳小蜂对扶桑绵粉蚧的寄生效能最高时的蜂蚧比为2:50,其差异显著;扶桑绵粉蚧密度过高或过低均会导致寄生率下降,原因可能是当班氏跳小蜂密度过大时,个体间的相互干扰导致寄生蜂搜索效能降低,而班氏跳小蜂密度过小时,则达不到寄生控制的效果,这与黄俊等(2012)的研究结果基本一致。组建了扶桑绵粉蚧第2、3代自然种群生命表,并分析了种群趋势指数(I)、干扰作用控制指数(IIPC)和排除作用控制指数(EIPC),结果表明,班氏跳小蜂对扶桑绵粉蚧具有良好的控制作用,放蜂区扶桑绵粉蚧第2、3代自然种群增长的数量最低分别下降到对照区的24.31%和12.82%,最高分别为34.79%和30.67%。运用排除作用控制指数(EIPC)分析表明,相比“捕食和其它”因子,班氏跳小蜂“寄生蜂寄生”因子是控制扶桑绵粉蚧种群数量发展的重要因子。由此可见,班氏跳小蜂是扶桑绵粉蚧重要的优势种天敌,其在扶桑绵粉蚧生物防治中的应用前景极为广阔。
陈华燕,何嫏芬,郑春红,李盼,易晴辉,许再福.2011.广东和海南扶桑绵粉蚧的天敌调查.环境昆虫学报,33(2):269-272.
何嫏芬,冯东东,李盼,许再福.2012.班氏跳小蜂对寄主龄期选择的研究.环境昆虫学报,34 (3):328-332.
黄俊,吕要斌,张娟,黄芳,贝亚维.2012.班氏跳小蜂对扶桑绵粉蚧的寄生功能反应.昆虫学报,55(12):418-1423.
黄玲,刘慧,肖铁光,周社文.2011.扶桑绵粉蚧寄生性天敌班氏跳小蜂生物学研究.应用昆虫学报,48(6):1734-1737.
马骏,胡学难,刘海军,梁帆,赵菊鹏,冯黎霞,陈乃中.2009.广州扶桑上发现扶桑绵粉蚧.植物检疫,23(2):35-36.
庞雄飞.2000.植物保护剂的评价方法.应用生态学报,11(1):9-14.
庞雄飞,梁广文.1995.害虫种群系统的控制.广州:广东科技出版社,22-24.
庞雄飞,梁广文,尤民生.1986.种群生命系统研究方法概述.天敌昆虫,8(3):176-186.
庞雄飞,梁广文,尤民生,吴伟坚.1988.昆虫种群生命系统研究的状态方程.华南农业大学学报,9(2):1-10.
庞雄飞,梁广文,曾玲.1984.昆虫天敌作用的评价.生态学报,1:46-56.
吴定发,李迎红,杨奇志,余冬林,熊况忠,曹婧,肖铁光.2011.扶桑绵粉蚧在中国的研究现状及其防治.作物研究,25(3):295-298.
武三安,张润志.2009.威胁棉花生产的外来入侵新害虫-扶桑绵粉蚧.昆虫知识,46(1):159-162.
朱艺勇,黄芳,吕要斌.2011.扶桑绵粉蚧生物学特性研究.昆虫学报,54(2):246-52.
Abbas G,Arif MJ,Saleem MS,Shah F.2012.Studies on biology ofPhenacoccussolenopsisTinsley(Hemiptera:Pseudococcidae)in Punjab,Pakistan -a pest of cotton and other crops.Pakistan Entomologist,34(2):131-136.
Akintola AJ,Ande AT.2008.First record ofPhenacoccussolenopsisTinsley(Hemiptera:Pseudococcidae)onHibiscusrosa-sinensisin Nigeria.Agricultural Journal,3(1):1-3.
Arif MJ,Gogi MD,Arshad M,Ashraf A,Suhail A,Zain-ul-Abdin,Wakil W,Nawaz A.2012.Host-plants mediated population dynamics of cotton mealybug,PhenacoccussolenopsisTinsley(Hemiptera:Pseudococcidae)and its parasitoid,AenasiusbambawaleiHayat(Hymenoptera:Encyrtidae).Pakistan Entomologist,34(2):179-184.
Dhawan AK,Singh K,Saini S,Aneja A,Singh J.2011.Parasitizing potential of parasitoid(Aenasiusbambawalei)on mealybug(Phenacoccussolenopsis)in cotton(Gossypium spp.)and weed plants.Indian Journal of Agricultural Sciences,81(1):97-99.
Fand BB ,Gautam RD,Suroshe SS.2011.Suitability of various stages of mealybug,Phenacoccussolenopsis(Homoptera:Pseudococcidae)for development and survival of the solitary endoparasitoid,Aenasiusbambawalei(Hymenoptera:Encyrtidae).Biocontrol Science and Technology,21(1):51-55.
Hayat M.2009.Description of a new species ofAenasiusWalker(Hymenoptera:Encyrtidae),parasitoid of the mealybug,PhenacoccussolenopsisTinsley(Homoptera:Pseudococcidae)in India.Biosystematica,3:21-26.
Hodgson C,Abbas G,Arief MJ,Saeed S,Karar H.2008.Phenacoccussolenopsis,an invasive mealybug damaging cotton in Pakistan and India,with a discussion on seasonal morphological variation.Zootaxa,1913:1-35.
Huang J,Lu YB,Zhang J,Huang F,Bei YW.2012.Parasitic functional response ofAenasiusbambawaleiHayat(Hymenoptera:Encyrtidae)toPhenacoccussolenopsisTinsley(Hemiptera:Pseudococcidae).Acta Entomologica Sinica,55(12):1418-1423.
Pala RV,Saini RK.2011.Biology ofAenasiusbambawaleiHayat(Hymenoptera:Encyrtidae).Journal of Insect Science,24(1):99-101
Tanwar RK,Jeyakumar P,Singh A,Jafri AA,Bambawale OM.2011.Survey for cotton mealybug,Phencoccussolenopsis(Tinsley)and its natural enemies.Journal of Environmental Biology,32(3):381-384.
