木地肤的开花动态与花粉活力及柱头可授性研究
2014-04-10郭红超严成魏岩
郭红超,严成,魏岩*
(1.新疆农业大学草业与环境科学学院 新疆草地资源与生态重点实验室,新疆 乌鲁木齐830052;2.中国科学院新疆生态与地理研究所,新疆 乌鲁木齐830011)
木地肤(Kochiaprostrata)为藜科(Chenopodiaceae)地肤属(Kochia)的多年生小半灌木,广布于欧亚大陆的干旱和半干旱地区,是中亚干旱地区适口性好、营养价值高的牧草品种,是牛羊爱吃的饲料之一[1-2],具有较高的耐盐和耐旱生长能力,对退化的盐化土壤具有改良的潜力[3]。此外,木地肤还可以用作植被恢复的物种[4]。作为一种分布范围广泛、生态价值较高、饲用价值较好的优良牧草,迄今为止国内外已有很多研究,但主要集中在形态[5]、营养器官的结构[6]、生态型[4]、饲用价值[7]、种子萌发[8-10]等方面;而对其花粉活力、柱头可授性等方面的研究相对较少。
系统地了解植物花的性表达特征和繁育特性,是研究植物生活史的前提,也是开展相关研究的基础[11]。适应传粉要求是花部进化的根本动力因素之一[12]。在有性生殖中,植物花的性表达特征与传粉者行为、传粉机制和植物适合度间有着内在的联系[13],是植物在传粉系统的选择压力下对环境适应的进化趋势。研究植物花的性表达特征对于深入理解植物繁殖生物学特性以及植物物种的形成具有重要意义。本文从开花动态、花粉活力、柱头可授性等角度对木地肤的开花和性表达特征进行研究,以揭示其繁殖生物学特性及其生态适应对策,为新疆荒漠地区饲用半灌木良种的繁育提供理论基础,为受损生态系统的保护、恢复提供依据。
1 材料与方法
1.1 材料与研究区自然概况
木地肤根粗壮,通常多分枝,具一年生短的不育枝和长的生殖枝,叶互生,腋生短枝常数片集聚呈簇状,花两性,通常2~3朵集生于叶腋,排列在当年生枝条上部呈穗状花序。
研究区位于新疆乌鲁木齐雅玛里克山,地处准噶尔盆地南缘,属于低山砾质荒漠,植被类型为小半灌木荒漠小蓬群系,天然植被以小蓬(Nanophytonerinaceum)、散枝猪毛菜(Salsolabrachiata)、紫翅猪毛菜(Salsolaaf-finis)、木地肤为主,盖度20%~30%。该地区属中温带荒漠区,春秋季较湿润,夏季干旱,热量充足,冬季漫长而寒冷[14]。
1.2 研究方法
1.2.1 开花动态的观察 野外观察于2013年4月中旬—10月中旬进行。分别在3个种群中选取10株健康、长势均匀的植株,定株观察木地肤的开花动态:随机选取10个穗状花序,挂牌标记。从现蕾开始,每天观察1次,直到花期结束。各花序中随机选取5朵单花,雄蕊散粉当天每隔0.5h观察1次,此后每天1次,直到果实成熟。每次观测时记录花蕾的变化、花序内花朵开放的顺序、花序开放持续时间,单花花朵开放、花被片伸展、雌雄蕊的变化。
1.2.2 花粉和胚珠比(P/O)的测定 选取将要开放的花20朵取下花药用1mol/L HCl水解去除药壁制成10 mL花粉粒悬浮液,吸取5μL悬浮液用血球计数板计数花粉数,重复10次,统计每朵花的花粉量,因为每朵花仅有1个胚珠,故其P/O值即为每朵花花粉数。依据Cruden[15]的标准,P/O为2.7~5.4时,其繁育系统为闭花受精;P/O为18.1~39.0时,繁育系统为专性自交;P/O为31.9~396.0时,繁育系统为兼性自交;P/O为244.7~2588.0时,繁育系统为兼性异交;P/O为2108.0~195525.0时,繁育系统为专性异交;亦即P/O值的降低代表近交程度的升高,P/O值的升高表明远交程度的上升[15]。
1.2.3 花粉活力的检测 用MTT法测定花粉的活力和寿命[16]。具体方法是:在盛花期将当日开放的花朵取回,置于室内;将散粉后不同时间(0,4,8,12,24,48,72h)花朵的花粉置于载玻片上,同时将少量新鲜花粉置于载玻片上用酒精灯加热杀死以用作对照;将1滴MTT蔗糖溶液(100mg MTT溶解在5mL 50g/L蔗糖溶液中)滴加到花粉样品中,充分混匀,让其风干后再重复置1滴MTT蔗糖溶液,干后在显微镜下观察并统计着色花粉粒和未着色花粉粒的数目(每片花粉数>500粒,重复10次,每次用3朵花)。若花粉变为蓝黑色则表明有活力,若变为黄褐色或无变化则表明无活力。

1.2.4 柱头形态特征观察与柱头可授性检测 借助显微镜,观察同一植株从柱头初露至散粉后不同时间阶段柱头的形态特征。用MTT法测定柱头的可授性[16]。具体方法是:在盛花期取柱头初露至散粉后不同时间阶段的柱头各20个,置于凹面载玻片的凹陷处,每个样品各加1~2滴 MTT蔗糖溶液(100mg MTT溶解在5mL 50 g/L蔗糖溶液中),充分浸透每个柱头,约1h后观察柱头的颜色变化。若柱头颜色呈深紫色,则显示柱头有活力,具有可授性;若柱头颜色不变,则显示柱头无活力,已失去可授性。
2 结果与分析
2.1 木地肤的开花物候与开花动态
观测结果表明,研究地区的木地肤花期为5月下旬到9月上旬,始花期为5月下旬,6月初进入盛花期,末花期8月初开始,整个花期为89d(5月26日至8月24日)。每个穗状花序的花朵全部开放需60~80d,单花花期为11~13d。
木地肤穗状花序的开花顺序为:中上部→下部→顶部,位于同一生殖枝中上部的花序先开放,大约5~6d后位于同一生殖枝枝条下部的花序开始大量开放,最后是枝条上部的花序开放;侧枝上的花序在主枝下部开花时开始开花,开花顺序为自下而上;对于同一聚头花序的小花而言,其开花顺序则为由内向外开放,即中间小花先于两边小花开放。
同一枝条上的木地肤柱头在雄蕊散粉前就已成熟并完成异花授粉,单花花期可分为雌性亚期和雄性亚期,雌性亚期为10~12d,雄性亚期为1d(图1)。从现蕾到柱头露出的时间为3~5d,柱头露出花被片,开花过程进入雌性亚期;根据柱头的形态变化可以将雌性亚期分为5个阶段:柱头露出、柱头展开、完全展开、授粉、萎蔫,柱头形状从卷曲到伸展再到干枯萎蔫;柱头完成授粉后,子房快速生长,小花在雌性亚期末期长大呈饱满的椭圆球形,之后花被片开裂,原来卷曲的雄蕊从花被片中伸展出,开花过程进入雄性亚期。雄蕊露出苞片时间一般在8:30-10:30,露出时间随温度和湿度有所变化,温度高时时间提前,湿度大时,时间推迟;雄蕊露出1h后,花被片外翻打开,此时有2~3个雄蕊的花丝伸直把花药推到柱头的外侧上方,约1h后剩余的花丝也伸直,托出花药,花药开裂,花粉散出。散粉持续3h左右,花药开裂后,残留花药壁不脱落,但颜色变暗褐色,散粉完毕后,花被片合拢,留有花药残壁的花丝合拢到花被片上方成束状,花药残壁于第2天脱落,花丝于第3天脱落。野外观察表明红色花药散粉时间一般比黄色花药晚30~60min。
2.2 木地肤的花粉和胚珠比(P/O)
木地肤的P/O值见表1。从表1中可以看出,木地肤红色和黄色花药的单花花粉量为(61900±2106)~(64200±2511),两者之间无显著差异(t=1.253,df=4,P>0.05);木地肤胚珠数为1枚,因此,木地肤的花粉/胚珠比(P/O)为 (61900±2106)~(64200±2511)(表1)。

表1 木地肤的花粉/胚珠Table 1 Pollen-ovule ratio of K.prostrata
2.3 木地肤的花粉活力和寿命
木地肤雄蕊有红、黄两种颜色花药,从表2可以看出在相同生境下,两种颜色花药刚散粉时(0h)花粉活力最高,达80%以上,72h后绝大部分花粉失去活力。相同生境下,不同颜色花药的花粉寿命有明显不同。黄花药和红花药的花粉活力在刚散粉时(0h)无差异(t=0.293,df=8,P>0.05),黄花药的花粉活力在雄蕊散粉8h内维持在60%以上,之后迅速降低至10%以下,即花粉寿命约为8h;红花药花粉活力在雄蕊散粉后逐渐降低,散粉4h内维持在65%以上,之后快速下降,8h后已下降到30%左右,花粉寿命约为4h。
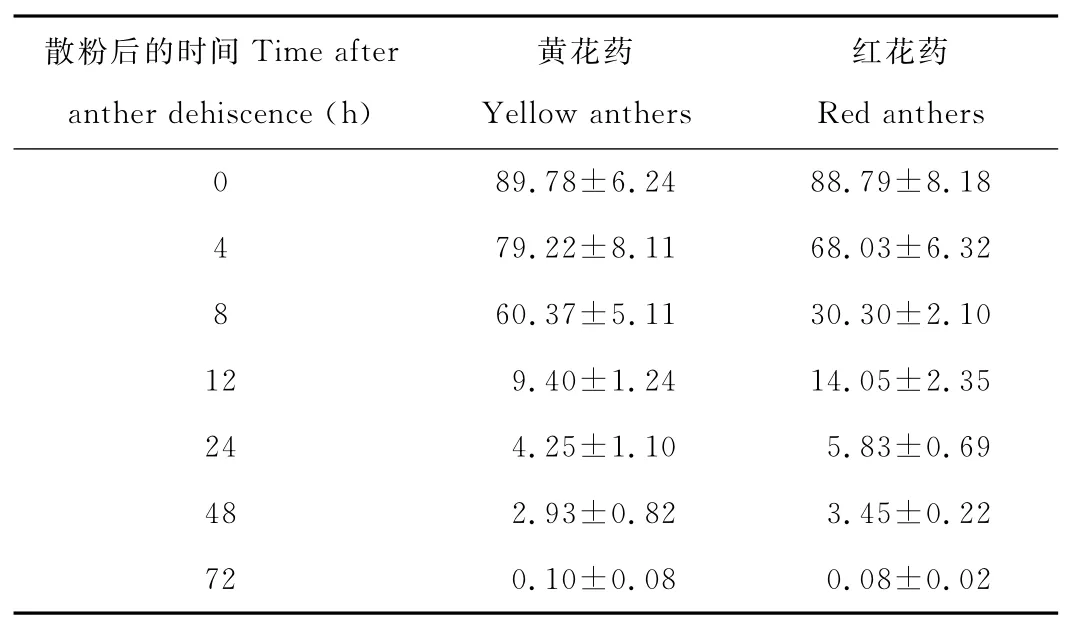
表2 木地肤的花粉活力Table 2 Pollen viability of K.prostrata %
2.4 柱头的形态特征及其可授性
木地肤有红黄两种颜色柱头,每种柱头在雄性亚期又有红黄两种颜色的花药,因此木地肤的花有4种不同的柱头花药组合类型,但以黄柱头/黄花药,黄柱头/红花药居多。
木地肤开花时柱头先从未展开的花被片中间伸出,此时柱头呈抱合球状,呈黄红色和嫩黄色,经检测有弱可授性(表3);6~7d后柱头逐渐展开,呈浅红色或黄色,显微镜下观察柱头乳突细胞明显生长,有分支出现,可以捕捉到花粉粒,可授性较强,可以进行授粉;再经过2~3d,柱头进入完全伸展期,此时的柱头呈双叉天线状,颜色为紫红色或黄色,柱头表面的乳突细胞最长,透明有活力(图2),经检测,此时的柱头可授性也最强,表现为刚加入MTT蔗糖溶液,柱头就变为紫黑色,柱头进入最佳授粉期;木地肤红色和黄色柱头的柱头可授性变化趋势是一致的,均在柱头露出6~7d后有较强的可授性;8~10d后可授性最强,柱头进入最佳授粉期,柱头颜色也变深;10~12d后,柱头完成授粉,乳突细胞失活脱水,柱头萎缩干枯,颜色变暗,逐渐丧失可授性;雄蕊传粉当天,花药散粉后,随着花被片的闭合,花柱枯萎,柱头可授性迅速丧失(表3)。值得注意的是,在雄蕊散粉前没有进行授粉的柱头一直保持强可授性,直到雄蕊散粉后花被片闭合时随花丝一起枯萎。
3 讨论
植物在长期进化过程中产生了各种花的发育机制以促进异交,并使自交以及雌雄功能之间的相互干扰维系在最低限度。同时,雌雄蕊产生了许多适应环境的机制,这些机制与雌雄功能、花被发育及环境之间是相互适应的[17-20]。
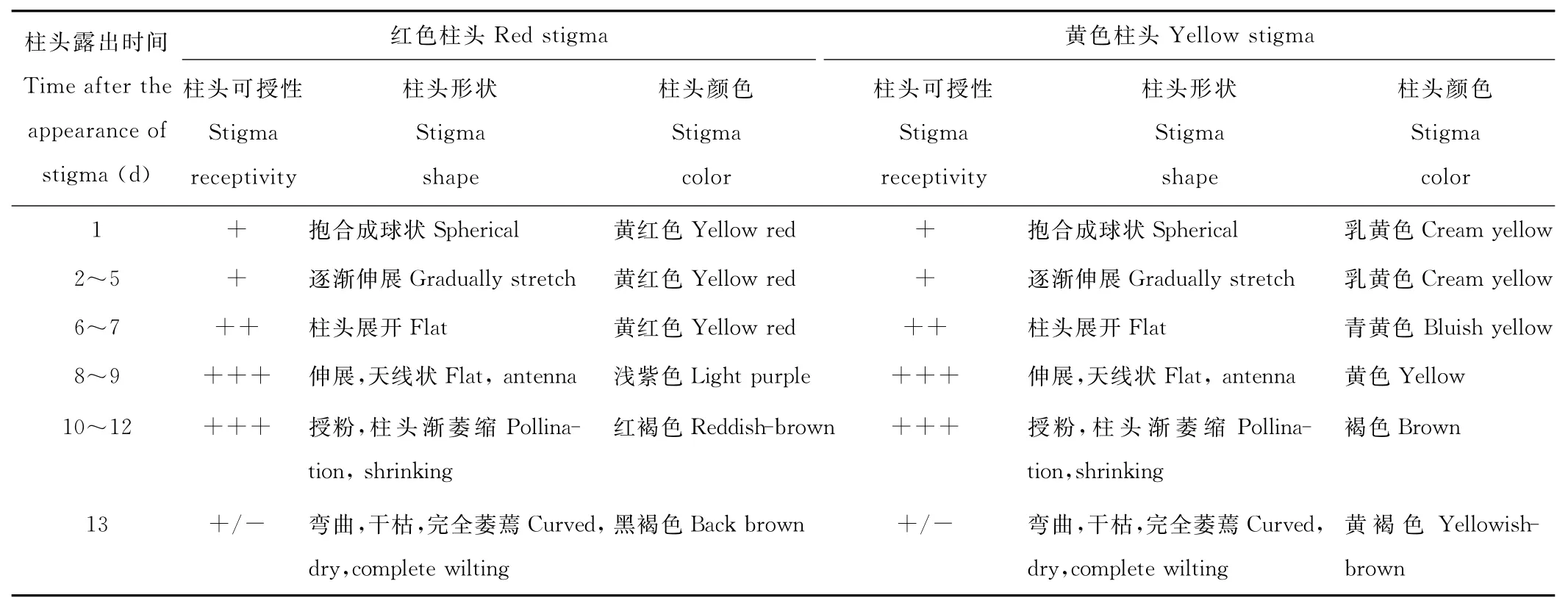
表3 木地肤两种颜色柱头的可授性及其形态特征Table 3 Stigma receptivity and its morphology of the stigma of different colors of K.prostrata

图1 木地肤开花进程组图(以黄柱头/红花药为例)Fig.1 Pictures of the flowering course of K.prostrata (The yellow stigma/red anther as an example)

图2 开花过程中柱头细胞及柱头的形态和颜色变化(红柱头)Fig.2 Changes in the shape and color of stigmas in the process of flowering(Red stigma)
Knuth[21]首次提出了雌雄异熟现象。按照两性功能出现的时间顺序雌雄异熟可分为雌性先熟(protogyny)和雄性先熟(protandry),但是雌性先熟在促进异交方面更为有效,因为雌性先熟植物的柱头成熟先于花药,可以优先接受异株植物花粉实现异交[22]。目前,发现在毛茛科、莎草科、木兰科、十字花科、车前科、桑科、樟科、睡莲科和玄参科等均存在雌性先熟现象[23-24]。风媒传粉的植物大多存在雌性先熟现象[22,25]。Bertin和 Newman[24]调查发现,88.5%的风媒植物是雌性先熟,而其他传粉方式的植物中只有34.8%具有雌性先熟。木地肤为小半灌木,属风媒花植物,具有一系列适应风媒传粉的生物学特征:花两性,小型,不鲜艳,无花冠;具大量的花粉,P/O值大;柱头呈双叉状,上有乳突细胞,便于捕捉花粉。木地肤在开花过程中具有雌性先熟的重要特征,柱头在刚露出苞片时就具有可授性,随着柱头不断长大变化,柱头可授性渐强,柱头可授期10~12d,最佳可授期3~4d,绝大部分柱头在这一期间完成异花授粉过程,授粉完成后,同一枝条上的雄蕊才露出花被片,花药裂开,并在一天内完成散粉过程。肾叶臭菘(Symplocarpusrenifolius)是具有雌性先熟的物种,单花花期为23.8d,其中雌性阶段为9.2d,占开花时间的38.7%,较长的雌性阶段为该物种增加异交机率提供了保证[26]。木地肤的单花花期为11~13d,其中雌性阶段为10~12d,占开花时间的92.3%,所以,木地肤存在的雌性先熟机制可以为异交的繁育系统提供良好的保证。
花粉保持生活力时间的长短,一方面由遗传基因所决定,另一方面也受环境因素的影响[27],如茄(Solanum melongena)的花粉在夏季只能存活1d,在冬季则能维持3d;海枣(Phoenixdactylifera)的花粉活力可维持数月至1年,是花粉寿命最长的植物[28],刺五加(Acanthopanaxsenticosus)长花丝植株的花粉活力比中花丝植株高,花粉活力及其下降快慢随生境不同而有所差异[29]。本研究中,观察到木地肤雄蕊在种群内有分化现象,有红、黄两种颜色的花药。两种花药的花粉活力在雄蕊散粉之初都较高(>80%),黄花药能持续8h之后迅速降低,红花药比黄花药晚开裂30~60min,只能维持4h,4h后花粉活力降到30%。植物花粉释放和传粉过程具有高度专一化和苛刻性[30]。短暂的花粉寿命使得木地肤必须具有高效的传粉机制、大量的花粉,以及多样化的传粉媒介才能保证传粉成功。木地肤两种颜色花药的高花粉活力以及开裂时间的分化一定意义上延长了花粉寿命,降低了雄性竞争,促进了花粉的输出和成功授粉率,是木地肤对恶劣环境的一种适应机制。
柱头可授性是有性生殖中最主要的环节之一,其可授期的长短直接影响花的授粉[31]。不同植物的柱头可授期所持续的时间从几小时到十几天不等,单花期的长短、开花后的天数、1d内的不同时辰以及柱头分泌物的有无等对其均有影响。如多枝柽柳(Tamarixramosissima)柱头在花开放之前就已具有较强可授性,开花散粉时到开花后约4h可授性最强[32],冷蒿(Artemisiafrigida)的柱头保持活力在6d左右,并以开花后第23天的活力达到最高[33]。另外,柱头的可授性与其形态特征之间也有密切的对应关系[34]。木地肤柱头有黄红两种颜色,但柱头可授性无差异。柱头可授期持续时间长达11~13d,最佳可授期长3~4d,柱头授粉后,柱头萎蔫,柱头乳突细胞失活,颜色变暗,两种柱头的颜色变化和柱头可授性都有一定的相关性:随着开花传粉过程,柱头颜色从浅到深再到暗黑,柱头可授性也相应从弱到强再到丧失。需要注意的是,雄蕊散粉当天个别未完成授粉的柱头仍保持强可授性,直到雄蕊散粉结束。这从一定意义上延长了柱头可授期的长度,更重要的是和自花散粉发生短暂重叠,时间上具备了自交的可能,是否存在延迟自交现象还需要通过人工套袋授粉实验和分子标记进一步证明。
Cruden[15]认为P/O值是两性花植物中表示性比的有效方法,能较好地反映繁育系统的基本类型。木地肤的P/O值为61900~64200,依据Cruden[15]的评判标准,其交配方式应属专性异交。一定的传粉效率是风媒植物种群进行有效繁殖的关键。在不利的传粉环境条件下,保障有性繁殖的资源投入,才能保障种群在恶劣环境中的遗传变异性和进化适应潜力[35]。从木地肤的性表达特征看,较高的P/O值、高花粉活力、长时间的柱头可授期以及雌性先熟特征为异花授粉提供了有力保障,使其在气候恶劣的准噶尔荒漠环境中有足够的花粉来进行有效传粉,是木地肤适应荒漠环境保证其生殖成功的有效对策。
[1] Nechaeva N T.Description of plants used for vegetative rang improvement[A].In:Nechaeva N T.Improvement of Desert Ranges in Soviet Central Asia[M].New York:Harwood Academic Publishers,1985:55-120.
[2] 冯缨,潘伯荣.新疆蒿类半灌木牧草资源分布及其饲用价值[J].干旱区资源与环境,2007,21(3):158-161.
[3] Balyan G A.Prostrate Summer Cypress and its Culture in Kirghizia[M].Isdatel’stvo,Frunze,Kirghizistan.Translated from Russian.1979.Published for the U.S.Dept.of Agriculture and the National Science Foundation by the Al Ahram Center for Scientific Translation,Springfield,VA.Available from National Tech.Information Service,1972:294.
[4] Acar R,Dursun D.Some features and important of forage kochia(Kochiaprostrata(L.)Schrad.)in natural areas of Konya[J].International Journal of Sustainable Water and Environmental Systems,2011,3(2):65-68.
[5] Shishkin B K.Flora of the USSR(Vol.6)[M].Moscow:Isdatel’stvo Akademii Nauk SSSR,1936.
[6] 李洪波,胡春元,贺晓,等.木地肤茎、叶解剖结构的观察[J].内蒙古农业大学学报,2008,2(4):85-88.
[7] Batanouny K H.Ecophysiology of halophytes and their traditional use in the Arab world[A].In:Choukr-Allah R,Malcolm C V,Hamdy A.Halophytes and Biosaline Agriculture[M].New York:Marcel Dekker Inc,1995:73-94.
[8] Waller S S,Britton C M,Schmidt D K,etal.Germination characteristics of two varieties ofKochiaprostrate[J].Journal of Range Management,1983,36(2):242-245.
[9] 刘涛,李柱,安沙舟.干旱胁迫对木地肤幼苗生理生化特性的影响[J].干旱区研究,2008,25(2):231-235.
[10] Orlovsky N S,Japakova U N,Shulgina I,etal.Comparative study of seed germination and growth ofKochiaprostrateandKochiascoparia(Chenopodiaceae)under salinity[J].Journal of Arid Environments,2011,75:532-537.
[11] 肖宜安,何平,李晓红.濒危植物长柄双花木的花部综合特征与繁育系统[J].植物生态学报,2004,28(3):333-340.
[12] Grant V,Grant K A.Flower Pollination in the Phlox Family[M].New York:Columbia University Press,1965.
[13] 刘乐乐,杨晓,高天鹏,等.毛茛状金莲花花部结构的吸引功能[J].草业学报,2013,22(3):190-195.
[14] 中国科学院新疆综合考察队,中国科学院植物研究所.新疆植被及其利用[M].北京:科学出版社,1978.
[15] Cruden R W.Pollen-ovule ratios:a conservative indicator of breeding systems in flowering plants[J].Evolution,1977,31:32-46.
[16] Rodriguez-Riano T,Dafni A.A new procedure to assess pollen viability[J].Sexual Plant Reproduction,2000,12(4):241-244.
[17] Darwin C.The Different Forms of Flowers on the Plants of the Same Species[M].London:Murray,1877.
[18] Barrett S C H,Jesson L K,Baker A M.The evolution and function of stylar polymorphisms in flowering plants[J].Annals of Botany,2000,85:253-265.
[19] Barrett S C H.The evolution of plant sexual diversity[J].Nature Review Genetics,2002,3:274-284.
[20] Barrett S C H.Flower diversity and plant mating strategies[A].McGraw-Hill Yearbook of Science & Technology[M].New York:McGraw Hill Inc,2004:110-113.
[21] Knuth P.Handbook of Flower Pollination(English translation)(3volumes)[M].London,Oxford:Clarendon Press,1906.
[22] Lloyd D G,Webb C J.The avoidance of interference between the presentation of pollen and stigamas in angiosperms.I.Dichogamy[J].New Zealand Journal of Botany,1986,24:135-162.
[23] Sargent R D,Otto S P.A phylogenetic analysis of pollination mode and the evolution of dichogamy in angiosperms[J].Evolution Ecology Research,2004,6:1183-1199.
[24] Bertin R I,Newman C M.Dichogamy in Angiosperms[J].The Botanical Review,1993,59:112-152.
[25] Cruden R W.Temporal dioecism:Systematic breadth,associated traints and temporal patterns[J].Botanical Gazette,1988,149:1-15.
[26] Wada N,Uemura S.Size-dependent flowering behavior and heat production of a sequential hermaphrodite,Symplocarpus renifolius(Araceae)[J].American Journal of Botany,2000,87:1489-1494.
[27] 胡适宜.被子植物胚胎学[M].北京:高等教育出版社,1982:20-60.
[28] 胡适宜.被子植物生殖生物学[M].北京:高等教育出版社,2005:143-147.
[29] 刘林德,张洪军,祝宁,等.刺五加花粉活力和柱头可授性的研究[J].植物研究,2001,21(3):375-379.
[30] 黄利春,金樑,张树振,等.蝶形花亚科植物花粉释放机制[J].草业学报,2013,22(6):305-314.
[31] 红雨,方海涛,那仁.濒危植物蒙古扁桃花粉活力和柱头可授性研究[J].广西植物,2006,26(6):589-591.
[32] 姬慧娟,尹林克,严成,等.多枝柽柳的开花动态及花粉活力和柱头可授性研究[J].西北农林科技大学学报(自然科学版),2009,37(5):114-118.
[33] 张雅荣,宛涛,蔡萍,等.冷蒿的开花动态与花粉活力及柱头可授性研究[J].中国草地学报,2012,34(1):108-112.
[34] 刘林德,张萍,张丽,等.锦带花的花粉活力、柱头可授性及传粉者的观察[J].西北植物学报,2004,24(8):1431-1434.
[35] 李冰,刘左军,赵志刚,等.海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响[J].草业学报,2013,22(1):10-19.
