海滨雀稗液泡膜H+-PPase(PvVP1)5′端的克隆和序列分析
2014-03-26宋辉南志标蔡小宁钟小仙顾洪如
宋辉,南志标*,蔡小宁,钟小仙,顾洪如
(1.草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020;2.南京晓庄学院生物化工与环境工程学院,江苏 南京211171;3.江苏省农业科学院畜牧研究所,江苏 南京210014)
焦磷酸酶(pyrophosphatase,PPase,EC3.6.1.1)是以焦磷酸为底物的水解酶,该酶广泛地存在于自然界中,参与合成糖类、核酸和蛋白质等多种代谢途径中所形成的焦磷酸的水解。PPase通常分为2类,一类是存在于细胞质和细胞器基质中的可溶性酶类,即无机焦磷酸酶(inorganic pyrophosphatase);另一类是与膜结合的不可溶性酶类,也就是膜结合焦磷酸酶(V-PPase)[1]。植物中主要存在3种质子泵(H+-ATPase),即位于细胞质膜上的P型质子泵(P-ATPase)、液泡膜上的V型质子泵(V-ATPase)和 H+-转运无机焦磷酸酶(H+-PPase)[2]。在盐胁迫条件下,植物V-H+-ATPase和V-H+-PPase一起为液泡膜Na+/H+逆向转运蛋白提供质子驱动力,将细胞质中过多的Na+区域化于液泡中,从而减轻Na+的毒害[3]。
20世纪50年代,Kunitz[4]首次从酵母中纯化到无机焦磷酸酶,随后,Karlsson[5]从甜菜(Betavulgaris)根的膜制剂中分离到钾激活的V-PPase。但是,直到1992年,Sarafian等[6]才从拟南芥(Arabidopsisthaliana)的cDNA文库中第一次克隆出完整的液泡膜H+-PPase的cDNA序列,命名为AVP1(M81892)。随后,研究者发现,大多数陆生植物H+-PPase cDNA含有2283~2319个核苷酸的开放阅读框架(ORF),编码大约761~722个氨基酸残基,计算出的分子量约为80~81kDa。另外,研究者比较了11种不同陆生植物的液泡膜H+-PPase氨基酸序列发现,它们的同源性高达86%~91%[7-8]。现在普遍认为,液泡膜H+-PPase是由单基因编码,它的过量表达很有可能会增加质子力而提高植物的耐盐性。异源表达拟南芥液泡膜H+-PPase恢复了盐敏感酵母突变体的耐盐性,盐芥的液泡膜TsVP转入陆地棉(Gossypiumhirsutum)后使其比同型野生型植株在150和250 mmol/L NaCl处理下耐受性增强[9-10]。席杰军等[11]和 Wu等[12]成功地从多浆旱生霸王(Zygophyllumxanthoxylum)中克隆了液泡膜H+-PPase并证实该基因对霸王响应盐碱和干旱胁迫具有重要作用。
海滨雀稗(Paspalumvaginatum)又名夏威夷草,隶属于禾本科雀稗属植物,原产于热带和亚热带滨海地带。海滨雀稗分蘖密度高,生长旺盛,具有较强的抗逆性和广泛适应性,耐盐碱的性能尤为出众,可以使用海水浇灌。因此,笔者认为,从海滨雀稗中克隆出相应的耐盐基因,然后将目的基因转入非盐生牧草中以提高其耐盐性是可行的。
1 材料与方法
1.1 材料
海滨雀稗于2011年6月份在温室中培养;大肠杆菌(Escheriachiacoli)DH5α;BU-SuperScript RT KIT、IPTG和X-Gal购自Biouniquer;pMDTM18-T Vector购自TaKaRa;Phusion High-Fidelity DNA Polymerase购自NEB(北京)有限公司;胶回收试剂盒,Marker购自北京天根生化;RNA提取试剂盒,加A反应液购自盖宁生物科技(北京)有限公司,引物合成和序列测定由南京金斯瑞生物科技有限公司完成。
1.2 海滨雀稗总RNA的提取
依据RNA pure超纯总RNA快速提取试剂盒的说明书进行。
1.3 海滨雀稗中间片段的克隆
将提取的总RNA使用BU-SuperScript RT KIT试剂盒反转录成cDNA。以反转录的cDNA为模板,P1:GGYGGDTCBTCHATGGCYCT,P2:AYTTCTTDGCRTTRTCCC为引物进行PCR。将目的片段回收纯化,送南京金斯瑞生物科技有限公司进行序列测定。
1.4 海滨雀稗PvVP15′RACE的克隆
按照Biouniquer的反转录试剂盒说明书操作步骤进行,略加修改,先合成5′-RACE-Ready cDNA。根据所获得的海滨雀稗液泡膜H+-PPase基因部分cDNA序列信息,利用Primer Premier 5.0软件设计特异性引物GSP1:5′-AGAGACCAACCGCCACACACAAGAACA-3′;再设计1个5′端巢式PCR引物 GSP2:5′-GAGCAGCACAAGACGATTCAGCA-3′。通过2次PCR获得5′RACE扩增产物,通过2%琼脂糖凝胶电泳分离PCR产物,切下条带提取纯化,用于后续实验。
1.5 序列分析
将5′RACE扩增得到的片段连接到pMD18-T载体上,转入大肠杆菌DH5α,挑取阳性克隆子,提交南京金斯瑞生物科技有限公司进行序列测定。测序结果应用DNAstar、ContigExpress、Clustal x、MEGA4.0和NCBI网站上的BLAST比对工具分析。
2 结果与分析
2.1 海滨雀稗RNA的提取和鉴定
提取海滨雀稗叶片的总RNA,经过1.5%琼脂糖凝胶电泳分离,可清晰观察到,28SrRNA与18S rRNA的条带亮度基本呈1∶1。结果显示,提取的海滨雀稗总RNA符合实验要求(图1)。
图1 海滨雀稗总RNA的提取与鉴定Fig.1 Extraction and identification of P.vaginatumRNA
2.2 海滨雀稗PvVP1中间片段的扩增
以海滨雀稗cDNA为模板,P1,P2为引物进行RT-PCR扩增。电泳显示在1391bp存在一条明显的亮带,命名为PvVP1(图2)。
2.3 海滨雀稗PvVP1 5′RACE的扩增
使用5′RACE巢式PCR,扩增产物于2%琼脂糖凝胶电泳进行检测。由图3可知,在500和750bp之间有一条明亮的条带。经测序可知,目标条带序列长697bp。
2.4 海滨雀稗PvVP1 5′RACE序列的分析
通过对海滨雀稗PvVP1中间片段和5′RACE扩增产物序列的测定,经序列拼接(图4),该片段ORF核苷酸序列长1605bp,共编码535个氨基酸残基。通过BLAST对比发现:海滨雀稗PvVP1的核苷酸序列与玉米(Zeamays)相应基因的核苷酸序列同源性达到93%,而与其他植物的相关序列同源性也达到79%以上,与玉米氨基酸的同源性高达99%。
图2 海滨雀稗PvVP1中间片段PCR扩增结果Fig.2 PCR amplification result of P.vaginatummiddle fragment

图3 巢式PCR电泳结果Fig.3 The nested PCR products

图4 核苷酸序列及所编码的氨基酸序列Fig.4 Nucleotide sequence and the deduced amino acid sequence
将通过推测得到的PvVP1氨基酸序列与其他植物的氨基酸进行多重比对发现(图5),它与其他植物的氨基酸的保守区域多达495个,而不保守区域为40个。其中,与大麦(Hordeumvulgare)和水稻(Oryzasativa)的相似性为98%,与小麦(Triticumaestivum)的相似性为97%,与卡拉草(Leptochloafusca)和高粱(Sorghumbicolor)的相似性为95%。
通过使用MEGA 4.0构建PvVP1的系统发育树显示(图6),海滨雀稗PvVP1与玉米的亲缘关系最近,与水稻的亲缘关系次之,与莱茵衣藻(Chlamydomonasreinhardtii)和盐藻(Dunaliellaviridis)的亲缘关系较远。

图5 PvVP1氨基酸序列与其他植物H+-PPase氨基酸序列的多重比对Fig.5 The alignment of the amino acid sequence of PvVP1and H+-PPase from other plants
3 讨论
植物在漫长的进化过程中,逐渐形成了通过减少盐分的摄入、增强盐分的外排或使Na+区域化进入液泡而避免盐害的胁迫[13]。目前,研究者主要利用以上3种机制来增强陆地棉[10]、水稻[14]、烟草[15](Nicotianatabacum)等经济作物的耐盐性,而在牧草方面的研究鲜见报道。近年来,随着草地退化的趋势日益严重,生态环境的恶化日渐突出,为了改善生态条件和提高盐碱地区城乡的绿化以及选育优良的牧草显得越来越重要[16-17]。我国的研究者在此方面做出了巨大的贡献,尤其是在结缕草[18](Zoysiajaponica)、象草[19](Pennisetumpurpureum)、狗牙根[20](Cynodondactylon)等方面的研究已具有一定的影响力,但是利用基因工程的手段研究海滨雀稗的报道较少。2009年,邹轶等[21]通过研究海盐胁迫对海滨雀稗生长的影响表明,海滨雀稗对盐胁迫有较强的抗性。正是基于此研究,笔者认为,海滨雀稗的较强耐盐性很有可能是由体内的某些基因的上调表达造成的。因此,本实验室试图利用分子生物学技术对海滨雀稗的耐盐基因进行挖掘,以期通过转基因技术将海滨雀稗良好的耐盐基因转化耐盐性较差的牧草中,最终培育出适合在盐碱地生长的牧草。
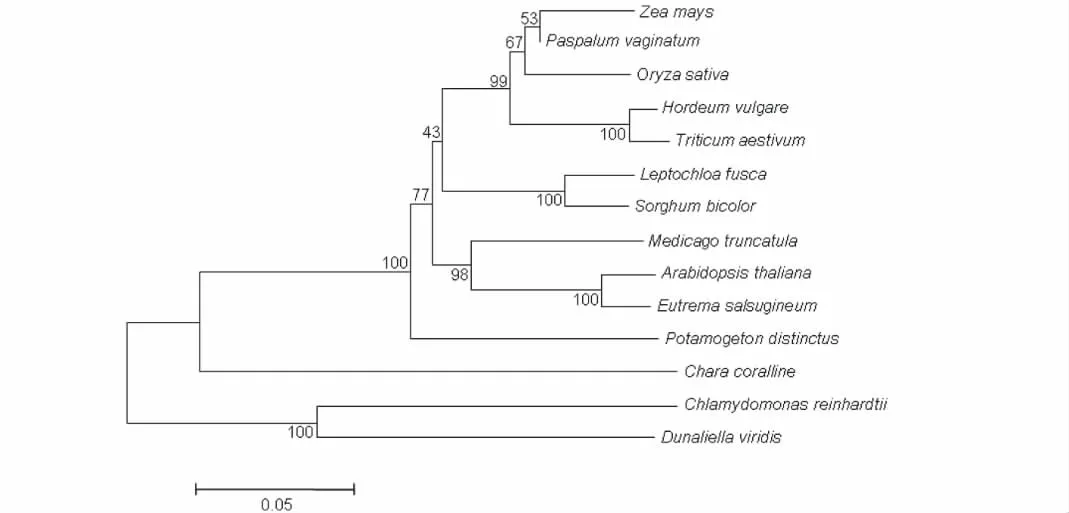
图6 海滨雀稗PvVP1的系统发育树Fig.6 The phylogenetic tree of PvVP1
目前普遍将液泡膜H+-PPase基因分为2种类型,即I型和II型[22-23]。在拟南芥中的研究发现,这2种类型的相似性只有36%,而且这2种类型的H+-PPase基因不是简单的同工酶关系,它们分布在液泡膜的不同部位,发挥不同的生理功能[24]。目前的研究表明,I型基因在植物的耐盐性中发挥重要的作用。本研究利用5′RACE技术快速克隆了海滨雀稗液泡膜H+-PPase基因,通过氨基酸比对发现,该基因与其他植物I型基因的氨基酸序列非常相似,说明所得到的克隆是液泡膜H+-PPase基因家族I类型。
在绿色植物和藻类植物的液泡膜H+-PPase基因中保守的GGG、KAADVGADLVGKVE、DNVGDNVGD、YYTS等基序在漫长的进化中一直未丢失,这被认为是液泡膜 H+-PPase酶的催化位点[7,25-26]。由于DVGADLVGKVE和DNVGDNVGD具有Gly(G)、Ala(A)、Asp(D)、Val(V)四种保守的氨基酸,因此这2种保守的基序不仅在进化上具有重要的意义;而且在液泡膜H+-PPase酶发挥生理功能时,也起着非常重要的作用[26]。本试验已成功地克隆到了上述4个保守的基序(图4),这表明海滨雀稗PvVP1具有液泡膜质子泵的功能。由前人的研究表明,液泡膜H+-PPase基因I型受 Mg2+和K+的激活[7],并且Asp(D)和Glu(G)被认为是最佳的金属离子螯合位点[27]。因此,笔者推测,DVGADLVGKVE和DNVGDNVGD这2个最为保守的基序最有可能是海滨雀稗PvVP1与Mg2+和K+的结合位点和发挥生理功能的催化位点。
通过以上对海滨雀稗PvVP1 5′末端序列的分析,笔者认为,海滨雀稗PvVP1具有良好的耐盐性。本实验室已准备下一步克隆海滨雀稗PvVP1的3′末端,从而克隆出PvVP1的全长序列用于转化优良牧草,以提高其耐盐性。
[1] 李小园,曾日中,校现周.植物焦磷酸的研究进展[J].生命科学,2004,8(4):83-87.
[2] Sze H,Schumacher K,Müller M L,etal.A simple nomenclature a complex proton pump:VHA genes encode the vacuolar H+-ATPase[J].Trends in Plant Science,2002,7(4):157-161.
[3] Zhu J K.Regulation of ion homeostasis under salt stress[J].Current Opinion in Plant Biology,2003,6:441-445.
[4] Kunitz M J.Crystalline inorganic pyrophosphatase inolated from baker’s yeast[J].Journal of General Psychology,1952,35:423-449.
[5] Karlsson J.Membrane-bound potassium and magnesium ion-stimulated inorganic pyrophosphatase form root and cotyledons of sugar beet(BetavugarisL.)[J].Biochimica et Biophysica Acta,1975,399:356-363.
[6] Sarafian V,Kim Y,Poole R J,etal.Molecular cloning and sequence of cDNA encoding the pyrophosphate-energized vacuolar membrane proton pump ofArabidopsisthaliana[J].Proceedings of the National Academy of Sciences,1992,89:1775-1779.
[7] Maeshima M.Tonoplast transporters:organization and function[J].Plant Molecular Biology,2001,52:469-497.
[8] Lv S Y,Jing Y X,Pang X B,etal.cDNA cloning of a vacuolar H+-pyrophosphatase and its expression in Hordeum brevisubulatum in response to salt stress[J].Agricultural Sciences in China,2005,4(4):247-251.
[9] Gaxiola R A,Rao R,Sherman A,etal.TheArabidopsisthalianaproton transporters,AtNhxlandAvp1,can function in cation detoxification in yeast[J].Proceedings of the National Academy of Sciences,1999,96:1480-1485.
[10] Lv S L,Zhang K W,Gao Q,etal.Over-expression of an H+-PPase gene fromThellungiellahalophilain cotton enhances salt tolerance and improves growth and photosynthetic performance[J].Plant and Cell Physiology,2008,49(8):1150-1164.
[11] 席杰军,伍国强,包爱科,等.多浆旱生植物霸王液泡膜 H+-PPase基因片段的克隆及序列分析[J].草业学报,2011,20(1):119-124.
[12] Wu G Q,Xi J J,Wang Q,etal.TheZxNHXgene encoding tonoplast Na+/H+antiporter from the xerophytesZygophyllumxanthoxylumplays important roles in response to salt and drought[J].Journal of Plant Physiology,2011,168:758-767.
[13] Zhu J K.Regulation of ion homeostasis under salt stress[J].Current Opinion in Plant Biology,2003,6(03):441-445.
[14] Carystinos G D,Macdonald H R,Monroy A F,etal.Vacuolar H+-translocating pyrophosphatase is induced by anoxia or chilling in seedlings of rice[J].Journal of Plant Physiology,1995,108:641-649.
[15] Gao F,Gao Q,Duan X G,etal.Cloning of an H+-PPase gene fromThellungiellahalophilaand its heterologous expression to improve tobacco salt tolerance[J].Journal of Experimental Botany,2006,57:3259-3270.
[16] 刘兴元,龙瑞军,尚占环.草地生态系统服务功能及其价值评估方法研究[J].草业学报,2011,20(2):167-174.
[17] 任继周,梁天刚,林慧龙,等.草地对全球气候变化的响应及其碳汇潜势研究[J].草业学报,2011,20(2):1-22.
[18] 李珊,陈静波,郭海林,等.结缕草属草坪草种质资源的耐盐性评价[J].草业学报,2012,21(4):43-51.
[19] 刘智微,钟小仙,常盼盼,等.海盐胁迫下苏牧2号象草幼苗不同器官中阳离子分配与运输[J].草业学报,2012,21(5):237-247.
[20] 陈静波,刘建秀.狗牙根抗盐性评价及抗盐机理研究进展[J].草业学报,2012,21(5):302-310.
[21] 邹轶,顾洪如,钟小仙,等.海盐胁迫对海滨雀稗生长及植株体内阳离子含量的影响[J].草业科学,2009,26(4):117-120.
[22] Maeshima M.Vacuolar H+-pyrophosphatase[J].Biochimica et Biophysica Acta,2000,1465:37-51.
[23] Gaxiola R A,Palmgren M G,Schumacher K.Plant proton pumps[J].FEBS Letters,2007,581:2204-2214.
[24] Drozdowicz Y M,Kissinger J C,Rea P A.AVP2,a sequence divergent,K+-insensitive H+-translocating inorganic pyrophosphatase fromArabidopsis[J].Journal of Plant Physiology,2000,123:353-362.
[25] Meng X Z,Xu Z H,Song R T.Molecular cloning and characterization of a vacuolar H+-pyrophosphatase fromDunaliella viridis[J].Molecular Biology Reports,2012,38:3375-3382.
[26] Hedlund J,Cantoni R,Baltscheffsky,etal.Analysis of ancient sequence motifs in the H+-PPase family[J].FEBS Journal,2006,273:5183-5193.
[27] Auld D S.Zinc coordination sphere in biochemical zinc sites[J].Biometals,2001,14:271-313.
