胡桃楸不同营养器官POD同工酶的动态变化
2014-03-22张丽敏封彦杰张卫东
张丽敏, 薛 晶, 封彦杰, 张卫东, 王 仲
(佳木斯大学 生命科学学院, 黑龙江 佳木斯 154007)
胡桃楸( Juglans Mandshurica Maxim)又名核桃楸、山核桃,为胡桃科胡桃属落叶乔木。主要分布于中国东北及华北地区,是东北地区著名的“三大硬阔树种”之一,为珍贵用材,也是著名的药用植物。同工酶是基因的直接产物,具有明显的种属、组织和发育阶段特异性。过氧化物酶(POD)同工酶是植物体中常见的氧化酶,广泛参与多种生理活动,植物体内的许多生理过程及植物的不同发育时间、环境条件的变化常与POD同工酶的种类及活性有关[1-3]。目前,有关胡桃楸研究的报道较多,但对胡桃楸POD同工酶的研究还未见报道[4-8]。本试验通过对胡桃楸不同器官POD同工酶进行生长期动态测定、比较分析,旨在掌握其同工酶谱带的变化规律,为更好地了解山核桃生长期特点,探讨山核桃生理代谢、发育特点与同工酶谱带的关系,为促进胡桃楸食药用开发利用、引种栽培提供生化水平参考。并试图为进一步探讨药用次生代谢产物的生成与POD活性大小及同工酶谱带数的消长关系,为探索山核桃药源不同生长发育期的特征提供相应的检测生化指标[1, 9-12]。
1 材料与方法
1.1 试验材料
胡桃楸整个生长周期可分为生长和休眠两个阶段,北方树种于5月初叶片开始萌发,10月末落叶,即5月—11月为胡桃楸的生长期,11月—次年4月为休眠期。生长期内生命活动旺盛,同工酶类、次生代谢产物等表达和积累丰富,故本供试材料胡桃楸取自黑龙江省佳木斯市四丰山生长结实较好的成片山核桃林,于2012年5月—11月,每月15日—20日取胡桃楸10棵优树混合得到树根、树皮(茎皮)、枝皮、树枝(当年生枝条)、树叶5个营养器官样品。依次用1,2,3,4,5编号。所采样品用湿布包裹,塑料袋封装,尽快带回实验室备用。
1.2 试验方法
1.2.1 样品制备
将胡桃楸各营养器官用清水洗净,再用蒸馏水漂洗,滤纸吸干后称重,每克鲜重加4 mL提取液(pH值7.2的蔗糖—磷酸缓冲液),冰浴下研磨至匀浆状,4层纱布过滤,将滤液3800 r/min离心15 min,上清液(样品)置冰箱(-20℃)备用。
1.2.2 电泳
采用聚丙烯酰胺凝胶垂直板电泳方法,分离胶浓度7.5%,浓缩胶浓度3.5%。4℃恒压(350 V)电泳,电极缓冲液为pH值 8.3 Tris-Gly系统。样品点样量均为15 μL,电泳重复2次。
1.2.3 染色及迁移率(Rf)的测定
采用醋酸联苯胺法染POD,待酶带显现清晰后,用流水漂洗后置蒸馏水中保存,Rf测定按照见参考文献[13]。
1.2.4 凝胶板的保存
用10%甘油浸泡玻璃纸8 h以上,然后在玻璃板上先放1张玻璃纸铺平,其上放凝胶,凝胶上再放1张玻璃纸,排除玻璃纸与凝胶间的气泡,阴干、保存,以备考证。
2 结果与分析
2.1 基本谱型
胡桃楸5个器官叠加后的酶谱型即本实验所观察到的胡桃楸POD同工酶基本谱型,共显现29条酶带,从负极到正极划分为A、B、C 3个区。A区Rf=0~0.47,共显现8条酶带A1至A8;B区Rf=0.48~0.70,共显现14条酶带B1至B14;C区Rf=0.71~1.0,共显现7条酶带C1至C7,依染色深浅区分为特强带、强带、次强带、弱带和痕迹带5个级别。
2.2 不同器官同工酶酶谱动态特征
2.2.1 根中POD同工酶酶谱变化
从电泳图谱(图1)可见,5月至11月,各生育阶段根中POD酶带变化明显。共显现15条酶带,A3、A5、A7、B1、B3、B7、B9、B10、C2、C4这10条酶带始终表达,为根部的基本谱带,其中A7、B1、C4在生长期内表达稳定,强度不变,而其余7条皆随生长期不同,酶表达强度发生变化,其差异可以作为表达各生长发育阶段系统发育差异的指标。其中B3、C2两条表现出5月、10月表达较强,其它月表达较弱的特点。5月天气渐暖,胡桃楸开始萌芽,此时期属于休眠转入生长期阶段,10月天气转寒,胡桃楸叶片脱落,此时期属于生长转入休眠期阶段。这两个时期都为胡桃楸生长期和休眠期的过渡时期,B3、C2在这两个时期表达较强可作为此过渡时期的特征性酶带。A3、A5、B7、B9、B10 5条随生长期延长,出现表达强度的周期变化。A2、B4、B11、B13、C3 5条酶带表达均表现出生长时期特异性,A2 、B4酶带出现5、6、7月表达,8、9月逐渐消失,10、11月又逐渐出现的周期性变化,B11、B13酶带5、6月表达,7月消失,8、9月出现,10月消失,11月再度出现,出现周期性变化。

图1 根POD同工酶
2.2.2 树皮(茎皮)中POD同工酶酶谱变化
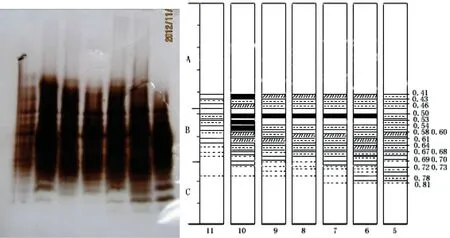
图2 茎皮POD同工酶
由电泳图谱(图2)可见5月至11月,茎皮中POD酶带变化明显,共显现18条酶带。A3、A5、A7、B1、B3、B9、B10、B11、C4这9条酶带皆表达,为茎皮部的基本谱带,但皆随生长期不同,酶表达强度发生变化,其差异可以作为表达各生长发育阶段系统发育差异的指标。酶带C4随生长期延长表达逐渐减弱为痕迹带,其余8条酶带均表现出11月表达较弱的特点且随生长期延长,表达强度表现周期性变化。另外B3酶带5、10月特异表达,可作为胡桃楸生长与休眠过渡时期的特异表达酶带。B8酶带7、8月特异表达,而此两月中B4、B12酶带消失,表现出生长期中生殖生长阶段特异性。此外茎皮的各酶带的表达皆出现11月表达强度下降或酶带消失的特点。北方胡桃楸10月落叶,11月进入冬季,胡桃楸已经进入休眠期,体内的各种生命活动,如:呼吸、蒸腾、芽的分化、根的吸收、养分合成和转化等,都进行得较微弱和缓慢,所以POD同工酶的表达也整体较弱。
2.2.3 枝皮中POD同工酶酶谱变化
从电泳图谱(图3)可见,6月至11月,枝皮中POD酶带变化明显,共显现17条酶带。B1、B3、B5、B9、B10、B11、B13这7条酶带始终表达,为枝皮部的基本谱带,其中B1、B3随生长期延长,酶带表达呈现由强渐弱的特点,B5、B9、B10、B11、B13 5条酶带均表现了8月出现表达强度高峰随后又下降的趋势。8月胡桃楸营养生长与生殖生长都达到了高峰期,POD同工酶作为植物生长期表达产生的一种重要的酶类,也从一个侧面体现出胡桃楸各阶段的生长发育特点。另外 A3、A5、A7、C2、C4 5条酶带6到10月较稳定表达,11月消失,可作为生长期特征性表达酶带。而A1、A2两条酶带从9至11月特异表达,11月也呈下降趋势。总体上茎皮的同工酶表达也出现11月表达强度下降或酶带消失的特点。
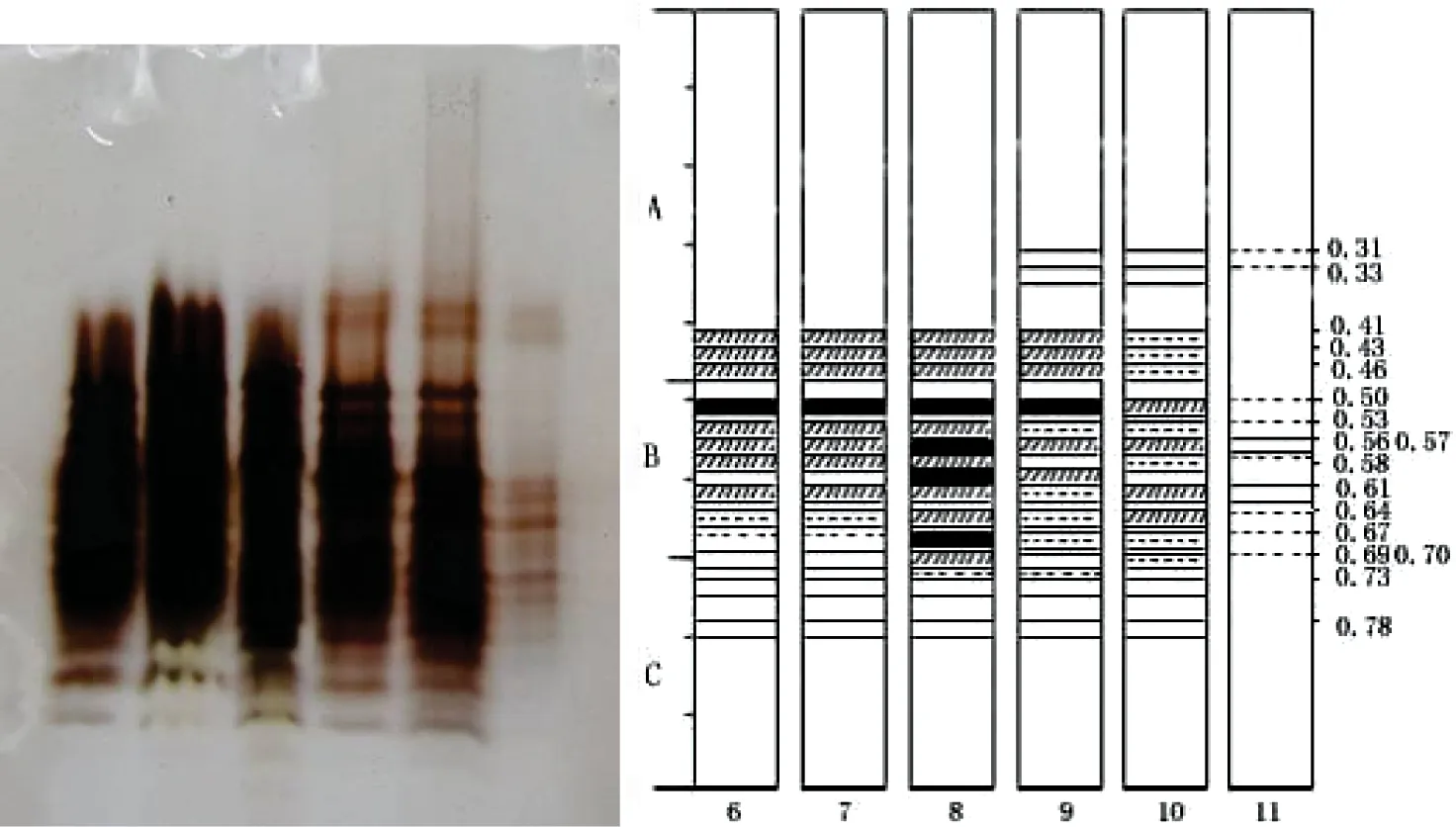
图3 枝皮POD同工酶
2.2.4 树枝中POD同工酶酶谱变化

图4 树枝POD同工酶
由电泳图谱(图4)可见,5月至9月,枝中仅显现7条酶带,变化较明显。A4、B1两条酶带始终表达,为枝部的基本谱带,且出现弱强弱的强度变化,均为6、7月表达强,A6、A8两条酶带5月不表达,B2酶带9月不表达,B5酶带5月、9月不表达,B7酶带8月特异表达。总体上枝中的同工酶表达5月酶带数量最少,其次是9月,而6、7、8月表达数量较多,活性较强。其表达特点与枝在这3个月中旺盛生长表现出一致性。
2.2.5 叶中POD同工酶酶谱变化
由电泳图谱(图5)可见,5月至9月,叶中POD酶带变化明显,共显现14条酶带。A4、B1、B2、B5、C2 5条酶带始终表达,为叶部的基本谱带,大致出现先弱后强再弱的强度变化。 A2、A3两条酶带9月不表达,A8、C3、C4酶带5月不表达,6月开始出现并渐强,A6酶带5月、9月不表达,C5、C7酶带8、9月特异表达,C6酶带7月出现并减弱。总体上叶中的同工酶表达5月酶带数量最少,而6、7、9月表达数量较多,8月表达数量最多,活性7、8月表达较强。可见POD同工酶在叶旺盛生长阶段表达种类多,活性强,与枝的POD同工酶表达特点相似。

图5 叶POD同工酶
2.3 不同器官酶谱叠加后的同工酶谱带特征
同工酶的基因表达存在着时间和空间的差异,为了更加准确地描述同工酶各酶带的显现情况,将每个月得到的各器官,根、茎皮、枝皮、枝、叶的酶谱叠加出一个总酶谱(叠加酶谱),可以直接观察到生长期中每月各器官中显现哪些酶带及共有酶带数(表1)。
5月POD同工酶4个器官共显现20条酶带,6月POD同工酶5个器官共显现25条酶带,7月POD同工酶5个器官共显现27条酶带,8月POD同工酶5个器官共显现27条酶带,9月POD同工酶5个器官共显现26条酶带,10月POD同工酶3个器官共显现19条酶带,11月POD同工酶3个器官共显现19条酶带,每个器官酶带数及共有酶带数见表1。
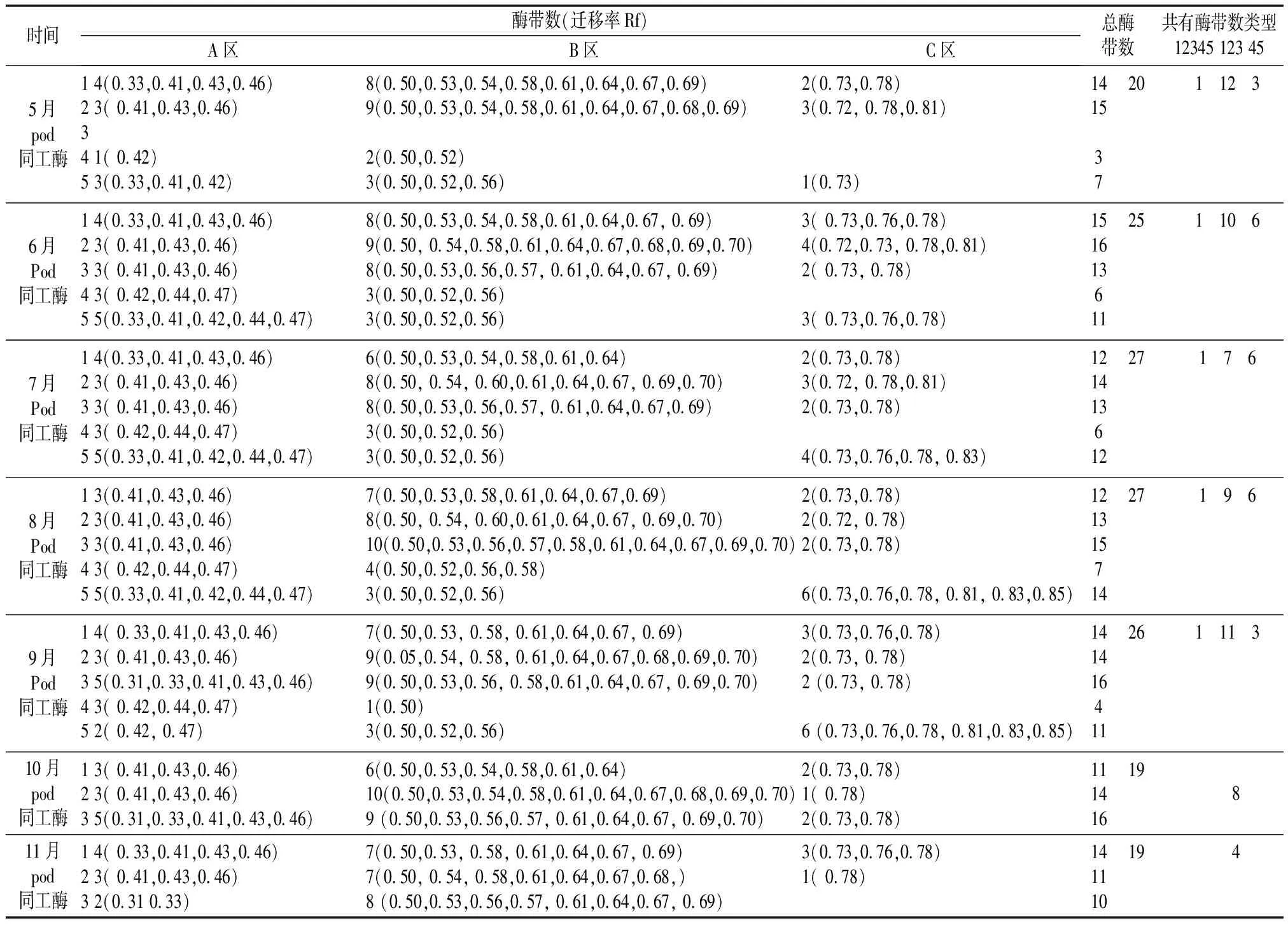
表1 不同生长期各器官叠加酶带特征
2.4 不同时期酶谱叠加后的同工酶谱带特征
将根、茎皮、枝皮、枝、叶5个器官的7个月份酶谱叠加出一个总酶谱(叠加酶谱),可以直接观察到各器官中每条酶谱带在生长期中是否显现以及显色强度(以显色最深时期为准)。根中5月至11月共显现15条酶带,茎皮中5月至11月共显现18条酶带,枝皮中6月至11月共显现17条酶带,枝中5月至9月共显现7条酶带,叶中5月至9月共显现14条酶带,各器官酶带具体分布情况及显色强度(表2)。
3 讨论
3.1 胡桃楸器官POD同工酶表达具有发育阶段性
通过胡桃楸根、茎皮、枝皮、枝、叶5个营养器官POD同工酶的测定分析,可以看出POD同工酶变化因生长时期的不同而不同,一方面,不同生长时期基本酶带有表达强弱的不同,生长期呈现动态变化,同时另一方面,特定时期某些酶带特异表达或消失,反映了代谢活动的不同。另外,从不同器官酶谱叠加后7个月的纵向比较,也可看出从萌芽期到落叶转入休眠期,表达的POD酶种类经历了由少增多再减少的周期性变化。可见POD可以作为一种重要酶类反映胡桃楸不同生长时期代谢活动的不同[1,14]。
3.2 胡桃楸POD酶带的共有性和同工酶的器官特异性[14]
将根、茎皮、枝皮、枝、叶的不同月份的电泳图谱叠加后可见既有共有酶带、也有各自的特有酶带(见表2)。5个器官电泳图谱中均有B1带,可能此酶带与胡桃楸的共同特性有关,或者说该酶带可能是胡桃属的特征性酶带。从叠加图谱中还可见到根中POD同工酶带数为15条,茎皮中为18条,枝皮中为17条,枝中为7条,叶中为14条,不论从酶谱带数目还是特有酶带上来看,胡桃楸POD同工酶表现出明显器官特异性。
3.3 胡桃楸pod同工酶分析宜采用根、茎皮、枝皮
胡桃楸5个器官中根部、茎皮部、枝皮部pod同工酶的种类多,活性强,酶带清晰稳定,故pod同工酶分析采用根、茎皮、枝皮部较合适。而枝中整个生长期酶带数较少(7条),活性较低;叶中pod同工酶带分离不清晰,故枝和叶不适合分析测定。

表2 各器官生长期叠加酶带特征
(其中+:有此酶带;-:无此酶带。)
(1级:特强带;2级:强带;3级:次强带;4级:弱带;5级:痕迹带)
参考文献:
[1]邹春静,盛晓峰,韩文卿.同工酶分析技术及其在植物研究中的应用[J].生态学杂志,2003,22(6):63-69.
[2]刘 稳,李 杨,高培基,等.过氧化物酶研究进展[J].纤维素科学与技术,2000, 8 (2): 50-64.
[3]吉志新,王 飞,王长青,等.蒙古沙潜不同发育时期酯酶和过氧化物酶同工酶的比较分析[J].河北科技师范学院学报, 2007, 21 (2): 19-22.
[4]崔 哲.胡桃楸的研究进展[J].现代医药卫生, 2008, 24 (20): 3061-3062.
[5]朱红波,赵 云,林士杰等.核桃楸资源研究进展[J].中国农学通报, 2011, 27 (25): 1-4.
[6]何惠芳.核桃楸青果皮研究现状[J].中医与中西医结合, 2011, 14 (12): 1834-1836.
[7]吕海宁,折改梅,吕 杨.核桃和核桃楸的化学成分及生物活性的研究进展[J].华西药学杂志, 2010, 25 (4): 489-493.
[8]许绍惠,唐婉屏,韩忠环.核桃楸毒性成分研究[J].沈阳农业大学学报, 1986, l7(2):34-39.
[9]周光宇,龚蓁蓁,王自芬.遗传育种的生化指标—同工酶分析[J].上海农业科技,1979(3):1-4.
[10]徐秀芳,张海洋,赵永勋,等.5个不同形态类型龙葵的同工酶研究[J].武汉植物学研究, 2001, 19 (1): 77-82.
[11]张东来,祁永会.黑龙江省胡桃楸用材林培育现状及前景[J]. 黑龙江生态工程职业学院学报, 2010, 23 (5): 18-19.
[12]陈翠英.核桃楸的经济价值与繁育技术[J].河北林业科技, 2008, (4): 49.
[13]张相歧.番茄属四个钟的POD同工酶的分析[J].植物研究, 1987, 7 (4): 131-136.
[14]雷泞菲,苏智先,陈劲松.同工酶技术在植物研究中的应用[J].四川师范学院学报:自然科学版 , 2000, 21 (4): 321-325.
