响应面法优化小刺青霉16-7产纤维素酶液体发酵工艺
2014-01-21薛桥丽李世俊胡永金
李 悦,薛桥丽,李世俊,王 晶,胡永金,*
响应面法优化小刺青霉16-7产纤维素酶液体发酵工艺
李 悦1,薛桥丽2,李世俊1,王 晶3,胡永金1,*
(1.云南农业大学食品科学技术学院,云南 昆明 650201;2.云南农业大学图书馆,云南 昆明 650201;3.玉溪沃森生物技术有限公司,云南 玉溪 653100)
在单因素试验的基础上,采用Plackett-Burman试验设计及响应面分析,利用Minitab软件,对纤维素酶高产菌小刺青霉(Penicillium spinulosum)16-7进行发酵工艺条件的优化。通过Plackett-Burman试验筛选出影响产酶的3 个主要因素,即稻草-麸皮(碳源)添加量、培养温度和培养时间。在此基础上通过最陡爬坡试验和响应面分析法进行回归分析。结果表明:当稻草-麸皮添加量为3.45 g/100 mL、培养温度为27.11 ℃和培养时间为146.27 h时,酶活力最高,此条件下滤纸酶酶活力预测值为132.53 U/mL。经过修正,选择稻草-麸皮添加量3.45 g/100 mL、培养温度27 ℃、培养时间146 h,此条件下测得羧甲基纤维素酶酶活力为387.58 U/mL、滤纸酶酶活力为128.86 U/mL,滤纸酶酶活力比优化前提高49.07%。
纤维素酶;发酵;Plackett-Bumann设计;响应面分析;小刺青霉
纤维素是地球上分布最广泛、含量最丰富的可再生资源,据不完全统计,全球每年通过光合作用产生的植物生物量高达1.14×1012t[1-2],其中大部分尚未被利用或未被合理利用,目前全世界被开发利用的农林纤维副产物不足2%,我国约有50%以上的农林废弃物在田间地头被白白烧掉。这不仅浪费了宝贵的自然资源,而且全世界每年因农林废弃物焚烧造成的直接经济损失达数十亿元[3-5]。
因此,合理开发和利用这一丰厚的天然资源——纤维素,是世界各国当前研究开发的重点领域[6-8]。如用纤维素酶将纤维素降解为可利用的糖液,再进一步转化为酒精、菌体蛋白、气体燃料(如氢气)等物质,对解决当今世界所面临的粮食短缺、能源危机和环境污染问题具有深远的意义[9-10]。目前用于生产纤维素酶的微生物菌种大多都是丝状真菌,主要为木霉属、曲霉属和青霉属等菌株[11]。
纤维素酶是微生物分解利用纤维素时自身合成的一种胞外酶,酶产量的高低直接影响纤维素利用率。在发酵过程中,纤维素酶的活力大小主要与菌种纤维素酶基因的表达有关,但是同一菌株,通过一定的外部条件优化,可以提高其产酶活力,发酵的工艺条件对产酶的影响也十分重要,以本实验室筛选保存的小刺青霉(Penicillium spinulosum)16-7为研究对象,设计Plackett-Burman试验,拟筛选出对纤维素酶发酵最重要的影响因素,结合响应面试验设计,优化纤维素酶液态发酵工艺条件。
1 材料与方法
1.1 菌种、试剂与培养基
1.1.1 菌种
小刺青霉(Penicillium spinulosum)16-7,云南省大围山原始森林土壤中筛选,由云南农业大学食品科学技术学院微生物实验室分离保存。
1.1.2 试剂
羧甲基纤维素钠(sodium carboxyl methyl cellulose,CMC-Na)(化学纯)、3,5-二硝基水杨酸(化学纯)国药集团化学试剂有限公司;酒石酸钾钠(分析纯)天津市永大化学试剂有限公司;定量滤纸(中速) 杭州富阳特种纸业有限公司。
1.1.3 培养基
种子培养基[11]:CMC-Na 1.5 g/100 mL、NH4NO30.1 g/100 mL、酵母膏0.1 g/100 mL、MgSO4·7H2O 0.05 g/100 mL、KH2PO40.1 g/100 mL,pH值自然,121 ℃灭菌30 min。
液体发酵培养基[11]:1.5 g/100 mL不同碳源、NH4NO30.4 g/100 mL、酵母膏0.4 g/100 mL、MgSO4·7H2O 0.02 g/100 mL、KH2PO40.4 g/100 mL,pH值自然,121 ℃灭菌30 min。
1.2 仪器与设备
HX121-0054立式压力蒸汽灭菌锅 上海华线医用核子仪器有限公司;SHA-BA恒温振荡器 常州澳华仪器有限公司;101-2A电热鼓风干燥箱 北京中兴伟业仪器有限公司;SW-CJ-2D超净工作台 苏州净化设备有限公司;7200型可见分光光度计 尤尼柯(上海)仪器有限公司;DT5-2型低速台式离心机 北京时代北利离心机有限公司。
1.3 稻草、麸皮预处理
将100 g稻草、麸皮分别置于500 mL的3 g/100 mL氢氧化钠溶液中混匀,0.15 MPa(126~128 ℃)保温5 min,放气[12]后备用。
1.4 培养方法
1.4.1 液体种子培养
将小刺青霉16-7制成孢子悬液(107~108个细胞/mL),按10%的接种量接入到装有100 mL液体种子培养基的250 mL三角瓶内,置于恒温摇床中28 ℃、120 r/min振荡培养3 d。
1.4.2 发酵培养
按照试验设计,将种子培养液接入发酵培养基进行发酵培养,每组做3 个平行样。
1.5 分析方法[13-15]
1.5.1 粗酶液的制备
将发酵液在5 000 r/min条件下离心10 min,取上清液作为粗酶液。
1.5.2 羧甲基纤维素酶活力(CMCase activity,CMCA)测定
分别向4 支试管(1 支空白管,3 支样品管)加入1 g/100 mL CMC-Na溶液(用pH 5.0醋酸-醋酸钠缓冲溶液配制)2.00 mL,然后加入一定稀释比例的酶液0.50 mL (空白管不加),混匀。将4 支试管置于50 ℃水浴中反应30 min,迅速向各管加入DNS试剂3.0 mL,再于空白管中加入稀释好的酶液0.5 mL。将4 支管同时放入沸水浴中,加热10 min取出。冷却至室温,定容至15 mL摇匀,以空白管(对照液)调零,测OD540nm值。酶活力单位采用国际定义方法:以1 mL酶液在1 min内分解底物生成1 μg葡萄糖的酶量定义为1 个酶活力单位(1 U/mL)。
1.5.3 滤纸酶活力(filter paper activity,FPA)的测定
分别向4 支试管(1 支空白管,3 支样品管),加入50 mg滤纸和pH 5.0醋酸-醋酸钠缓冲溶液1.5 mL,然后加入一定稀释比例的酶液0.50 mL(空白管不加),使管内溶液浸没滤纸,盖塞。将4 支试管同时置于50 ℃水浴中反应60 min后,立即向各管加入DNS试剂3.0 mL。再于空白管中加入稀释好的酶液0.50 mL。后续操作同1.5.2节。
1.6 试验设计
1.6.1 单因素试验
分别选取碳源、氮源、起始pH值、装液量、转速、接种量、培养时间、培养温度、表面活性剂添加量进行单因素试验,研究各因素对发酵产纤维素酶的影响。
碳源:首先分别以3 g/100 mL稻草、玉米秸秆、滤纸、葡萄糖、CMC-Na为不同的碳源,然后在确定稻草为最佳碳源基础上添加不同质量比例的麸皮,确定二者的添加比例以提高碳源利用率;氮源:以0.6 g/100 mL蛋白胨、酵母膏、NH4NO3、NaNO3、(NH4)2SO4及NH4Cl作为不同的氮源,确定最佳氮源后考察不同添加量对产酶的影响;培养基起始pH值:分别设培养基起始pH值为3.0、3.5、4.0、4.5、5.0、5.5、6.0、6.5;装液量:分别盛装25、50、75、100、125、150 mL液体培养基到250 mL的三角瓶中;摇床转速:分别设置为100、125、150、175、200、250 r/min;接种量:向产酶培养基中分别加入1%、2%、3%、4%、5%、6%、7%、8%、9%、10%的种子培养液;培养温度:在26、28、30、32、34 ℃条件下培养;培养时间:分别培养3、4、5、6、7、8、9 d;表面活性剂的添加量:向培养基中分别添加0.0、0.2、0.4、0.6、0.8、1.0、1.2 mL/100 mL的吐温-80。
1.6.2 Plackett-Burman试验
Plackett-Burman设计是一种两水平的试验设计方法,它基于非完全平衡块原理,可以用最少试验次数估计出因素的主效应,适用于从众多的考察因素中快速有效地筛选出最为重要的几个因素[16-17]。在单因素试验的基础上,以FPA作为响应值,对影响产酶的碳源、氮源、菌种接种量、摇床转速、装液量、吐温-80添加量、初始培养温度、初始pH值、培养时间9 个影响因素进行筛选,筛选出3 个主效应因子,每个因素选两个水平,另设3 个虚拟列,以考察实验误差,共20 组试验。
1.6.3 最陡爬坡试验
最陡爬坡试验可以达到快速进入最优点附近区域的目的[18]。系统最优条件的初步估计常常远离实际的最优点,要先采用最陡爬坡试验设计逼近最佳值区域后才能建立有效的响应面拟合方程[19-21]。以Plackett-Burman试验的零水平作为最陡爬坡试验的基础点,根据Plackett-Burman试验获得的3 个主效应因素的比例关系设定步长和上升路径进行最陡爬坡试验设计。
1.6.4 响应面试验设计
采用Box-Behnken法,依据Plackett-Burman试验和最陡爬坡试验确定的试验因素与水平,对纤维素酶发酵条件进行响应面分析试验,包括12 个析因试验和5 个中心试验,共17 组试验。
2 结果与分析
2.1 单因素试验
2.1.1 碳源对产纤维素酶的影响

图1 不同碳源对小刺青霉16-7产纤维素酶的影响Fig.1 Effect of carbon sources on cellulase production
由图1可知,小刺青霉16-7在以稻草为碳源时酶活力最高(CMCA为244.67 U/mL、FPA为69.55 U/mL),秸秆次之。说明该菌对稻草这类天然纤维素的分解利用能力较强,其中葡萄糖几乎是所有真菌都喜欢利用的良好碳源,所以酶活力也较高,但产酶能力仍低于天然纤维素,这和很多研究报道的葡萄糖是真菌纤维素酶合成的阻遏物的观点一致。结果说明,该菌在不同种类的碳源培养基中,产酶能力有很大差别。

图2 稻草与麸皮质量比对小刺青霉16-7产纤维素酶的影响Fig.2 Effect of rice straw-to-wheat bran ratio on cellulase production
由图2可知,随着稻草与麸皮比例的不断变化,酶活力呈现先增大后减小的趋势。当稻草与麸皮比为7∶3时,酶活力达到峰值,CMCA为269.83 U/mL,FPA为74.15 U/mL。分析原因可能是麦麸为产酶提供少量的淀粉、粗蛋白、微量元素等营养因子,从而提高菌株的产酶能力;同时,培养基内的初始糖质量浓度可加速诱导菌株产酶,且在一定范围内,诱导纤维素酶产量是呈正比关系,但如果糖质量浓度过高则会降低酶产量。由此可见,培养基中添加适当比例的麸皮时,对于诱导纤维素酶的形成是有帮助的。但是,随着麸皮的添加比例不断增大,会对纤维素酶的形成产生抑制作用,另外过多的麸皮可能会造成培养基的蓬松度降低,通气性减弱,从而影响纤维素酶的形成。

图3 稻草-麸皮添加量对小刺青霉16-7产纤维素酶的影响Fig.3 Effect of carbon source concentration on cellulase production
由图3可知,随着稻草-麸皮添加量的不断增大,酶活力呈现先急剧增大,到峰值后缓慢下降的趋势。当稻草-麸皮添加量为3 g/100 mL时,CMCA和FPA达到峰值,CMCA为281.77 U/mL,FPA为77.09 U/mL。可能是因为适量的稻草-麸皮添加量作为一种诱导剂在表达体系中起作用,过低的糖浓度诱导作用小,延缓产酶期,且菌丝体生长缓慢,而过高的糖质量浓度则会对纤维素酶的形成产生抑制作用。初步确定稻草-麸皮的添加量为3 g/100 mL。
2.1.2 氮源对产纤维素酶的影响

图4 不同氮源对小刺青霉16-7产纤维素酶的影响Fig.4 Effect of nitrogen sources on cellulase production
由图4可知,不同氮源对小刺青霉16-7产酶影响的差异较大,其中蛋白胨和酵母膏对产酶有明显的促进作用,其次为(NH4)2SO4、NH4Cl。结果表明,氮源对酶活力的影响因素依次为:有机氮>铵盐>硝酸盐,采用蛋白胨作为氮源时效果最好,CMCA为258.59 U/mL,FPA为69.28 U/mL。

图5 蛋白胨添加量对小刺青霉16-7产纤维素酶的影响Fig.5 Effect of peptone concentration on cellulase production
由图5可知,随着蛋白胨添加量的不断增大,酶活力呈现先增大后减小的趋势。当蛋白胨添加量为0.8 g/100 mL时,CMCA和FPA达到峰值,CMCA为282.32 U/mL,FPA为74.29 U/mL。初步确定蛋白胨的添加量为0.8 g/100 mL。
2.1.3 摇床转速对产纤维素酶的影响

图6 摇床转速对小刺青霉16-7产纤维素酶的影响Fig.6 Effect of rotation speed on cellulase production
由图6可知,当摇床转速为150 r/min时,酶活力均达到峰值,CMCA为275.41 U/mL,FPA为72.48 U/mL。当摇床转速从100 r/min增至150 r/min时,小刺青霉16-7纤维素酶酶活力呈增大趋势;但当转速超过150 r/min后,产酶水平有所下降,选择使用150 r/min的转速比较适合。
摇床转速主要影响发酵液的溶解氧浓度,目的菌是好氧菌,在生长和产酶过程中要消耗大量的氧,所以在产酶过程中应不断振荡来增加发酵液的溶氧浓度,另外适度振荡还可以增加菌丝与发酵液中营养物质的充分接触,使菌种能够获得充足的营养。由以上结果可以看出,转速太低可能导致培养基中溶氧不足,影响菌体生长;太高可能由于机械剪切力过大,造成菌丝断裂,进而影响产酶。
2.1.4 培养时间对产纤维素酶的影响

图7 培养时间对小刺青霉16-7产纤维素酶的影响Fig.7 Effect of culture time on cellulase production
由图7可知,培养时间对产酶有较大的影响。当培养时间为3~6 d时,酶活力呈现急剧增大的趋势;当培养时间为7 d时,酶活力达到顶峰,CMCA为270.68 U/mL,FPA为71.23 U/mL;当培养时间超过7 d时,酶活力呈现缓慢下降的趋势。
酶的产生需要一个过程,目的菌16-7培养前期处于迟缓期,菌丝体缓慢生长,对环境逐步适应,培养一段时间之后,菌丝体进入对数生长期,菌丝体生长旺盛,随后进入稳定期,是菌株产生代谢产物的主要阶段,产酶量逐步达到最高,这时的培养时间是7 d左右,即目的菌产纤维素酶的最佳培养时期。发酵后期,随着培养基内可供菌株利用的营养物质的减少,以及代谢产物的积累,菌体步入衰亡期,细胞相继衰老、死亡,生长繁殖等代谢活动减弱。通过对发酵过程中菌株生长状况的观察,发现菌丝体的形态变化与其产酶变化是一致的。
2.1.5 培养温度对产纤维素酶的影响
由图8可知,培养温度对产酶活力具有较大影响。在28 ℃条件下,CMC酶活力和滤纸酶活力均达到最大值,CMCA为270.48 U/mL,FPA为70.92 U/mL。当温度超过28 ℃,酶活力下降明显。初步确定目的菌产纤维素酶培养温度为28 ℃。

图8 培养温度对小刺青霉16-7产纤维素酶的影响Fig.8 Effect of culture temperature on cellulase production
在较低温度26 ℃时,菌株生长缓慢,传代时间延长,酶活力高峰期出现较晚。当培养温度为28 ℃时,菌丝体旺盛生长,产酶高峰期提前,酶分泌增多,酶活力增大。但若温度持续升高,可能造成了菌株过早老化,酶分泌量减少,酶活力逐渐降低。使得参与生化反应的酶系统遭到破坏,直接导致微生物细胞因无法繁殖而死亡。
2.1.6 培养基初始pH值对产纤维素酶的影响

图9 培养基初始pH值对小刺青霉16-7产纤维素酶的影响Fig.9 Effect of initial pH on cellulase production
由图9可知,培养基初始pH值对产酶有较明显的影响,当初始pH值在3.0~5.5时,随着pH值升高,纤维素酶活性出现上升趋势;当初始pH值为5.5时,酶活力最高,CMCA为286.54 U/mL,FPA为75.41 U/mL;当初始pH值大于5.5后,酶活力逐步下降。初步确定目的菌产纤维素酶初始pH值为5.5。这与大多数真菌适宜在中性偏酸性的环境中生长相符。
2.1.7 接种量对产纤维素酶的影响

图10 接种量对小刺青霉16-7产纤维素酶的影响Fig.10 Effect of inoculum size on cellulase production
由图10可知,接种量对产酶效果影响较为明显,接种量为1%~4%时,酶活力较小;当到5%时,产酶水平达到峰值,CMCA为294.35 U/mL,FPA为77.46 U/mL;超过5%以后,产酶水平有所下降,而在6%~8%,产酶较为稳定;当接种量大于8%,酶活力迅速下降。
接种量过小时,营养过剩,菌丝体剧烈生长,产酶滞后,产酶活力不高,延长生长周期;接种量过大时,第一,影响产酶培养基的透气性,从而使得酶活力降低;第二,发酵前期菌株生长迅速,培养基温度升高,抑制产酶能力;其三,菌株生长过密,发酵后期营养供给不足,竞争增强,抑制菌株生长。
2.1.8 装液量对产纤维素酶的影响

图11 装液量对小刺青霉16-7产纤维素酶的影响Fig. 11 Effect of liquid medium volume in 250-mL flasks on cellulase production
由图11可知,250 mL三角瓶装100 mL培养基能获得较佳的产酶效果,CMCA为289.56 U/mL,FPA为76.20 U/mL。装的过多和过少都不利于产酶,装液量过少,可能会引起摇瓶中的液体培养基晃动厉害,破坏了菌体与纤维素的黏附状态,且机械剪切力过大,造成菌丝断裂,进而影响产酶,但随着装液量超过100 mL,产酶水平也逐渐下降,这可能是由于供氧不足且在液体搅动过程中菌体不能和营养物质充分接触,不利于目的菌的生长。
2.1.9 吐温-80添加量对产纤维素酶的影响

图12 吐温-80添加量对小刺青霉16-7产纤维素酶的影响Fig.12 Effect of Tween-80 addition on cellulase production
由图12可知,吐温-80对产酶能力的影响较小。添加表面活性剂吐温-80能在一定程度上提高纤维素酶活力,添加量为0.6 mL/100 mL时酶活力达到峰值,CMCA为281.45 U/mL,FPA为74.07 U/mL。CMCA是未添加时的1.21 倍,滤纸酶酶活力是未添加时的1.23倍,再增加用量酶活力开始下降,这可能是因为表面活性剂加入过多可能会引起蛋白质变性,进而使酶活力降低。同样,添加量过低则起不到作用,添加量为0.2 mL/100 mL时的酶活力与未添加时相差不大。因此,添加表面活性剂控制在合适范围内,将有利于提高产酶能力。针对小刺青霉株16-7,本实验采用0.6 mL/100 mL的吐温-80添加量较为合适。
2.2 Plackett-Burman试验

表1 Plackett-Burman试验设计及结果Table 1 Plackett-Burman experimental design and results

表2 Plackett-Burman试验统计分析Table 2 Statistical analysis of the Plaekett-Burman experimental results
Plackett-Burman试验设计与结果如表1所示,统计分析见表2,满足“Prob>F”小于0.05的3 个因素(碳源、培养温度、培养时间)是显著的,可考虑作为主要因素进一步做响应面试验。而其他因素的取值则根据各因素效应的正负和大小,正效应因素均取较高值,负效应因素均取较低值,即选择蛋白胨添加量1.2 g/100 mL、摇床转速150 r/min、pH 5.5、装液量100 mL/250 mL、接种量7.5%、吐温-80添加量为0.6 mL/100 mL。方差分析见表3,主效应“Prob>F”等于0.012,说明模型与试验值拟合良好。

表3 Plackett- Burman试验方差分析Table 3 Variance analysis of the Plaekett-Burman experimental results
2.3 最陡爬坡试验
根据Plackett-Burman试验获得的3 个主效因素的比例关系设定步长和上升路径,其中培养温度有显著正效应,应增加;稻草-麸皮添加量和培养时间有显著负效应,应减小。进行最陡爬坡试验,试验设计及结果见表4。最优条件出现在试验2,因此以该处理条件为中心点进行下一步响应面优化试验。

表4 最陡爬坡试验设计及结果Table 4 The response values of steepest ascent path
2.4 响应面试验设计及结果
2.4.1 模型的建立
以FPA为响应值,根据表4的结果拟定试验因素水平与结果见表5,用Design Expert软件进行多元回归分析。经回归拟合后,试验因素对响应值的影响可以用以下回归方程表示:Y=132.25-2.03A+1.44B+1.99C+ 0.75AB-2.20AC+1.13BC-10.48A2-6.64B2-12.38C2。

表5 Box-Benhnken试验设计与结果Table 5 Box-Benhnken Experimental design and results

表6 响应面二次回归模型方差分析Table 6 ANOVA for the response surface quadratic model
由表6可知,所选模型的不同处理间差异极显著(P<0.000 1),说明用回归方程描述各因素与响应值之间的关系时,其应变量与全体自变量之间的线性关系显著,即该试验方法可靠。失拟项P=0.143 8>0.05,差异不显著,说明试验的误差等偶然因素不会对试验结果带来显著影响,该方程对试验拟合情况好,试验误差小。
该方程的决定系数R2=98.85%,说明模型可以解释98.85%的试验所得的产酶变化,表示方程拟合度良好。变异系数表示试验的精确度,变异系数值越低则试验的可靠性越高,本试验变异系数=1.35%,仅有1.35%的响应值变异不能用该模型解释,说明试验操作可信。
2.4.2 响应面图和等高线图分析
模型的三维响应面图和等高线图能比较直观地解释各变量和变量之间对响应值的影响。根据上述回归方程绘制出响应面图和等高图,见图13~15。利用该图可以将其3 个变量中的1 个变量固定在中心值水平,而对另外2 个变量交互影响FPA的效应进行分析与评价。

图13 碳源添加量和培养温度对纤维素酶产量影响的响应等高线图和响应面图Fig.13 Response surface and contour plots for the effects of carbon source concentration and culture temperature on cellulase production
由图13可知,在培养时间保持固定的情况下,稻草-麸皮添加量(A)和培养温度(B)对FPA的影响。等高图近圆形,且等高线较疏,表明此两因素的交互作用不显著。响应面立体分析图开口向下,当响应值增大到极值以后,随着各因素的增大,响应值逐渐减小。当稻草-麸皮添加量在3.25~3.75 g/100 mL、培养温度在26.5~27.5 ℃时,可得到FPA的最大值。

图14 碳源添加量和培养时间对纤维素酶产量影响的响应等高线图和响应面图Fig.14 Response surface and contour plots for the effects of carbon source concentration and culture time on cellulase production
由图14可知,在培养温度保持固定的情况下,稻草-麸皮添加量(A)和培养时间(C)对FPA的影响。等高图呈椭圆形,且等高线较密,可以直观地看出两因素的交互作用显著,同样从方差分析可知稻草-麸皮添加量(A)和培养温度(B)交互作用显著(P=0.028 1)。处于同一曲线上的FPA是一样的。在椭圆区域的中心FPA最大,由中心向边缘FPA逐渐降低。响应面立体分析图开口向下,当响应值增大到极值以后,随着各因素的增大,响应值逐渐减小。当稻草-麸皮添加量在3.25~3.75 g/100 mL、培养时间在132~156 h时,可得到FPA的最大值。
由图15可知,在稻草-麸皮添加量保持固定的情况下,培养温度(B)和培养时间(C)对FPA的影响。等高图呈椭圆圆形,且等高线较密,但方差分析中两因素的P=0.201 9,可以得出此培养温度和培养时间的存在一定交互作用。处于同一曲线上的FPA是一样的。在椭圆区域的中心FPA最大,由中心向边缘FPA逐渐降低。响应面立体分析图开口向下,当响应值增大到极值以后,随着各因素的增大,响应值逐渐减小。当培养温度在26.5~27.5 ℃、培养时间在132~156 h时,可得到FPA的最大值。
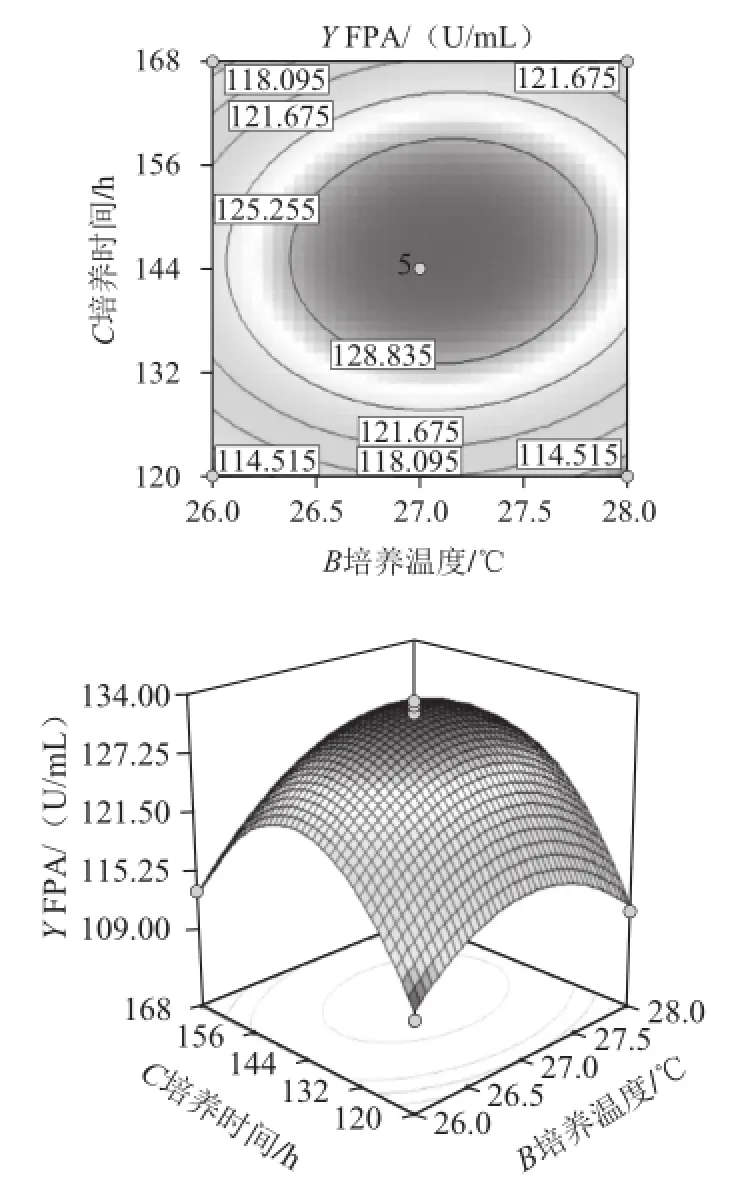
图15 培养温度和培养时间对纤维素酶产量影响的响应等高线图和响应面图Fig.15 Response surface and contour plots for the effects of culture time and temperature on cellulase production
对模型方程进行典型性分析,结果表明该模型有稳定点,最大值就是稳定点。此回归模型得出Box-Behnken试验的最优结果为稻草-麸皮添加量3.45 g/100 mL、培养温度27.11 ℃、培养时间146.27 h,此条件下FPA为132.53 U/mL。
经过修正,选择稻草-麸皮添加量3.45 g/100 mL、培养温度27 ℃、培养时间146 h,进行3 批次发酵验证实验,获得平均FPA为128.86 U/mL ,比优化前平均FPA(86.44 U/mL)提高49.07%。结果与模型预测基本一致,说明模型能够比较真实地反映各显著因素对纤维素酶产量的影响,表明所得模型对实验具有指导意义。
3 结 论
单因素试验的基础上,通过Plackett-Burman试验有效地找出影响纤维素酶产量主要因素,最陡爬坡试验能充分接近最大响应面区域,再利用Box-Behnken试验对显著因素进行优化。得到的发酵最优条件为:稻草-麸皮添加量3.45 g/100 mL、培养温度27.00 ℃、培养时间146.00 h、蛋白胨添加量1.2 g/100 mL、摇床转速150 r/min、pH 5.5、装液量100 mL/250 mL、接种量7.5%、吐温-80添加量为0.6 mL/100 mL。此条件下测得羧甲基纤维素酶酶活力为387.58 U/mL、滤纸酶酶活力为128.86 U/mL,滤纸酶酶活力比优化前提高49.07%。
参考文献:
[1] 陈洪章, 李佐虎. 纤维素原料微生物与生物量全利用[J]. 化工科技市场, 2001(5): 17-20.
[2] 邓天福, 杜开书, 李广领. 纤维素酶及其在酿造业中的应用[J]. 中国酿造, 2011 , 30(12): 17-19.
[3] SHULER M L. Utilization and recycle of agriculture wastes and residues[M]. Boca Raton, Florida: CRC Press Inc., 1980.
[4] 薛桥丽, 王炜, 胡永金, 等. 堆肥中高温纤维素酶菌株的筛选及其酶性质研究[J]. 中国酿造, 2012, 31(1): 30-33.
[5] 武林芝. 纤维素酶以及应用行业的研究进展[J]. 安徽农业科学, 2013, 41(26): 10570-10574.
[6] 宋颖琦, 刘睿倩, 杨谦, 等. 纤维素降解菌的筛选及其降解特性的研究[J]. 哈尔滨工业大学学报, 2002, 34(2): 197-200.
[7] 武香玉, 徐海燕, 辛国芹, 等. 纤维素酶及其研究进展[J]. 饲料博览, 2013(10): 27-29.
[8] 胡格华, 苏香萍, 潘虹, 等. 纤维素酶产生菌的筛选及产酶条件的研究[J]. 三峡大学学报: 自然科学版, 2013, 35(4): 100-102.
[9] LUTZEN N W, NIELSEN M H, OXENBOELL K M, et a1. Cellulase and their applications in the conversion of lignocellu-loses to fermentable sugars[J]. Philosophical Transactions of the Royal Society, 1983, 300: 283-291.
[10] 傅力, 涂正东, 叶凯. 里氏木霉高产纤维素酶菌株的选育及产酶培养基的优化[J]. 食品与机械, 2009, 25(3): 10-13.
[11] PERSON I, TJERNELD F, HAHN-HǍGERDAL B. Fungal cellulolytic enzyme production: a review[J]. Process Biochemistry, 1991, 26: 65-74.
[12] 叶生梅, 许盼吉, 汪江林. 稻草秸秆的预处理及发酵乙醇的试验研究[J]. 中国粮油学报, 2008, 35(3): 16-20.
[13] 刘海波, 王义强, 陈介南. 一株高产纤维素酶菌的筛选与鉴定[J]. 生物学杂志, 2008, 25(3): 16-20.
[14] 张瑞萍. 纤维素酶的滤纸酶活和CMC酶活的测定[J]. 印染助剂, 2002, 19(5): 51-53.
[15] MANDELS M, ANEREOTTI R E. Enzymatic hydrolysis of waste cellulose[J]. Biotechnology and Bioengineering, 1976, 6: 17-21.
[16] ZHOU Jiangya, YU Xiaojuan, DING Cong, et a1. Optimization of phenol degradation by Candida tropicalis Z-04 using Plackett-Burman design and response surface methodology[J]. Journal of Environmental Sciences, 2011, 23(1): 22-30.
[17] 潘春梅, 王辉, 任敏. 响应面法优化L-谷氨酰胺发酵培养基的研究[J].生物技术通讯, 2013, 19(4): 528-531.
[18] CHEN Xiaochun, BAI Jianxin, CAO Jiaming, et a1. Medium optimization for the production of cyclicadenosine 3,5-monophosphate by Mierobacterium sp. No.205 using response surface methodology[J]. Bioresource Technology, 2009, 100(2): 919-924.
[19] RATNAM B V V, RAO M D. Optimization of fermentation condtions for the production of ethanol from sago starch using response surface methodology[J]. World Journal of Microbiology and Biotechnology, 2003, 19: 523-526.
[20] BAS D, BOYAEL I H. Modeling and optimization I: usability of response surface methodology[J]. Journal of Food Engineering, 2007, 78: 836-845.
[21] FUNG W Y, WOO Y P, LIONG M T. Optimization of growth of Lactobacillus acidophilus FTCC 09291 and evaluation of growth characteristics in soy whey medium: a response surface methodology approach[J]. Journal of Agricultural and Food Chemistry, 2008. 56: 7910-7918.
Optimization of Cellulase Production by Penicillium spinulosum 16-7 in Liquid-State Fermentation by Response Surface Methodology
LI Yue1, XUE Qiao-li2, LI Shi-jun1, WANG Jing3, HU Yong-jin1,*
(1. College of Food Science and Technology, Yunnan Agricultural University, Kunming 650201, China; 2. Library of Yunnan Agricultural University, Kunming 650201, China; 3. Walvax Biotechnology Co. Ltd., Yuxi 653100, China)
This paper reports the optimization of the fermentation conditions for cellulase production by Penicillium spinulosum 16-7 by Plackett-Burman design and response surface methodology using the statistical software Minitab. By using Plackett-Burman design, carbon source (a combination of rice straw and wheat bran at a mass ratio of 7:3) concentration, culture temperature and time were identified as main factors that influence cellulase production. The selected variables were further investigated by steepest ascent analysis and optimized by response surface analysis. The highest cellulase activity was obtained after culture at 27.11 ℃ for 146.27 h at a carbon source concentration of 3.45 g/100 mL, leading to a predicted filter paper activity of 132.53 U/mL. Under the modified fermentation conditions of 3.45 g/100 mL carbon source concentration, 27 ℃ and 146 h, carboxymethyl cellulase activity was 387.58 U/mL and the filter paper activity was 128.86 U/mL, 49.07% higher than that obtained before optimization.
cellulase enzyme; fermentation; Plackett-Bumann design; response surface analysis; Penicillium spinulosum
Q93.33
A
1002-6630(2014)17-0137-09
10.7506/spkx1002-6630-201417028
2014-04-16
国家自然科学基金地区科学基金项目(30960308);云南省自然科学基金项目(2008ZC057M)
李悦(1988—),女,硕士,主要从事食品质量及安全性研究。E-mail:xixiliyue@tom.com
*通信作者:胡永金(1972—),男,教授,博士,主要从事功能性食品与生物技术研究。E-mail:huyjin@126.com
