玉树典型嵩草草甸植物及土壤特性初探
2013-04-25孙长宏
孙长宏
(称多县草原站,青海 称多 815100)
探究高寒嵩草(Kobresia)草甸退化成因一直都是众多学者研究的热点之一[1]。关于高寒嵩草草甸的退化机理主要集中在几个方面。“超载放牧”理论[1]认为,放牧家畜的选择性采食引起植物群落优势种的演替是高寒嵩草草甸退化的主要原因[2]。因此,通过围封轮牧能达到治理高寒嵩草草甸退化的目的。马玉寿等[3]研究指出,对于轻度、中度退化草地,一定年限的围栏封育后草地可以得到恢复,然而,对于重度退化草地和“黑土型”极度退化草地,围封的方式很难使嵩草草甸恢复到初始状态[3]。针对该问题,曹广民等[1]提出了“高寒嵩草草甸的被动-主动”退化假说。可以说该理论是对“超载放牧”理论的补充,该理论强调在重度、极度退化阶段,由于杂草的大量侵入,高寒嵩草草甸的被动退化转而成为主动退化,常规的围栏封育不能使退化草地恢复正常。因此,对于重度退化的嵩草草甸,人工建植、引入优良牧草成为治理重度退化嵩草草甸的唯一途径。虽然人工建植能短期内实现退化草甸的恢复,但并不能长期维持较高的生产能力和稳定的群落结构[4]。可见,关于高寒嵩草草甸的退化机理研究并未完全透彻。鉴于此,本研究试图从健康与退化嵩草草甸的植被性状和土壤因子的比较分析中探究影响高寒嵩草优良牧草生物量的主导因子,区分各环境因子的贡献程度,以期为退化嵩草草甸的修复提供科学参考。
1 材料和方法
1.1研究区概况 研究区位于青海省玉树县境内,95°41′40″~97°44′34″ E,33°44′44″~33°46′44″ N,属典型的高原高寒气候。全年无四季之分,仅有冷暖季之别,冷季7~8个月,暖季4~5个月。年均温2.9 ℃,1月均温-7.5 ℃,7月均温12.5 ℃。年降水量487 mm。地形以高原为主,西北和中部高,东南与东北部低。草地类型有嵩草草甸,金露梅(Potentillafruticosa)灌丛草甸等,但主要以嵩草草甸为主。主要优势种是小嵩草(K.pygmaea)、矮嵩草(K.humilis)、线叶嵩草(K.capllifolia)、藏嵩草(K.tibetica)等,主要伴生种有异针茅(Stipaaliena)、圆穗蓼(Polygonummacrophyllum)、珠牙寥(P.viviparum)等,并有龙胆(Gentianaspp.)、星状风毛菊(Saussureastella)等杂类草。主要土壤类型是高山草甸土、高山灌丛草甸土及沼泽土。
1.2试验方法
1.2.1样地设置 试验地位于玉树县境内巴塘乡的国营牧场当地农户冬季放牧草地。根据于健龙等[5]对健康与退化草甸的划分依据,选取当地的小嵩草草甸(KG)、重度退化小嵩草草甸(HD-KG)、藏嵩草草甸(KT)和重度退化的藏嵩草草甸(HD-KT)为试验样地。
1.2.2鼠洞调查 于2011年7月20日,分别在不同草地类型划分面积为100 m×100 m的样地,在不同样地内选取5个25 m×25 m且立地条件相似的样方,然后对每个样地进行堵洞填埋,并调查总洞口数和有效鼠洞。第1天将所有鼠洞口计数作为总洞口数,然后填埋,后3 d连续每天在15:00-17:00调查记录被高原鼠兔(Ochotonacurzoniac)刨开的新洞口并计数,然后重新填埋,每天被高原鼠兔刨开的新洞口即为当天的有效洞口,最后确定的每个样地的有效鼠洞数是连续3 d的平均值。
1.2.3样方调查 于2011年7月20日,在不同样地内随机选取5个10 m×10 m的样方,用直尺测量样方内裸地的最长、最宽处,换算成面积,记为裸地面积。同时,在样方内随机选取3个1 m×1 m的样方,记录样方内的物种数,并分别用直尺量取5株不同种的植物的株高,用针刺法测定植被盖度。在10 m×10 m的大样方内,随机用土钻钻取土壤,直至土钻扎到土壤基岩,取出土钻,观察土壤剖面,用直尺量取生草层厚度,重复5次。在植物生长期,采用收获法,用剪刀齐地面剪去植被地上部,按照莎草类、禾草类(优良牧草)和杂类草分类,分别称取优良牧草鲜质量、杂类草鲜质量后,在105 ℃烘干至质量恒定,莎草类和禾草类质量记为优良牧草质量,总质量记为地上生物量。
在测定地上生物量的同时,采用环刀法,在样方内不分层采集0~30 cm地下生物量,5次重复,自然风干后,过0.5 mm细筛,用细纱布包好根系,清水冲洗,并拣除石块和其他杂物,在105 ℃的烘箱内烘至质量恒定,即为地下生物量。用烘干法测定土壤质量含水量,重复3次,即为土壤含水量。在样方内采集植被根际0~15和15~30 cm的土壤样品,重复3次,混合后风干待测。其中土壤全氮用半微量凯氏定氮法测定,土壤全磷用钼锑抗显色法测定,土壤有机质用硫酸重铬酸钾法。
1.2.4数据分析 用Excel 2007软件作图,用SPSS 13.0软件进行方差、线性回归、相关分析和主成分分析。
2 结果与分析
2.1植被特征 鼠洞密度、裸地面积和植被高度均为退化小嵩草草甸显著高于小嵩草草甸,退化藏嵩草草甸裸地面积和植被高度显著大于藏嵩草草甸(P<0.05)(表1),但不同样地的物种数差异不显著。藏嵩草草甸的土壤含水量显著高于退化藏嵩草草甸(P<0.05),生草层厚度呈现相似的变化趋势(表1)。
2.2地上和地下生物量 小嵩草草甸的地上生物量、优良牧草质量和地下生物量都显著高于退化小嵩草草甸(P<0.05),藏嵩草草甸的优良牧草质量和地下生物量都显著高于退化藏嵩草草甸(P<0.05)(图1)。
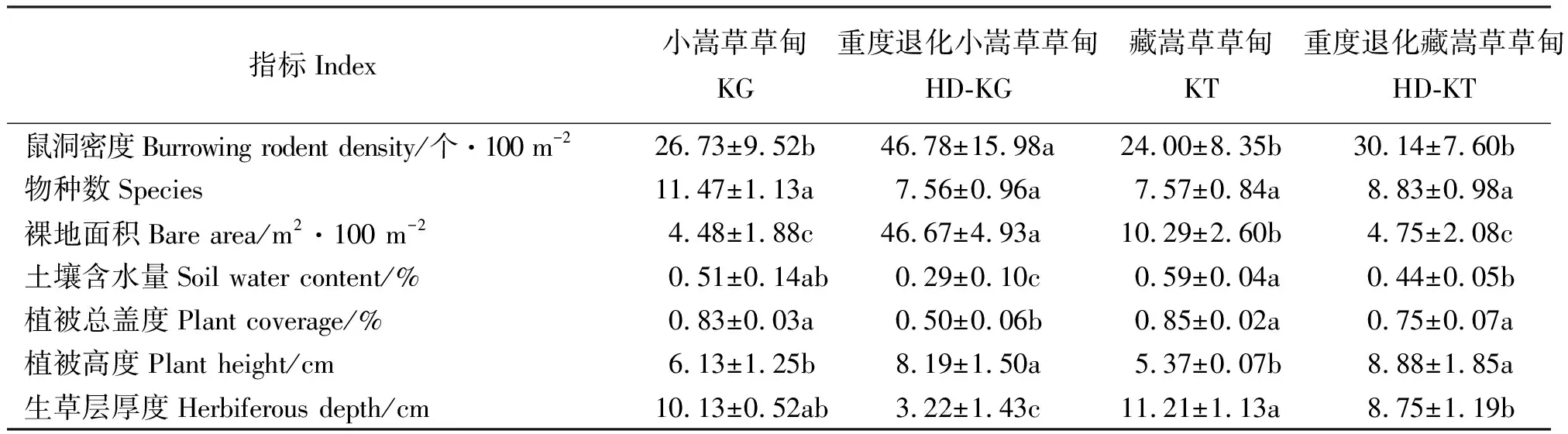
表1 不同样地样方指标Table 1 Plot indexes of different sites
2.3土壤养分 藏嵩草草甸的土壤全氮和有机质分别为11.48和312.26 g·kg-1,都显著高于退化藏嵩草草甸(P<0.05)。小嵩草草草甸的土壤全氮和有机质分别为5.69和114.69 g·kg-1,均显著高于退化小嵩草草甸(P<0.05)。不同样地的土壤全磷则差异不显著(图2)。
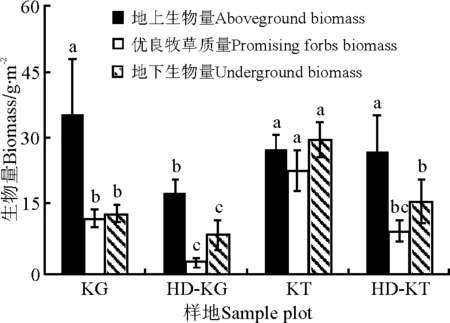
图1 不同样地生物量Fig.1 Biomass of different sample plots
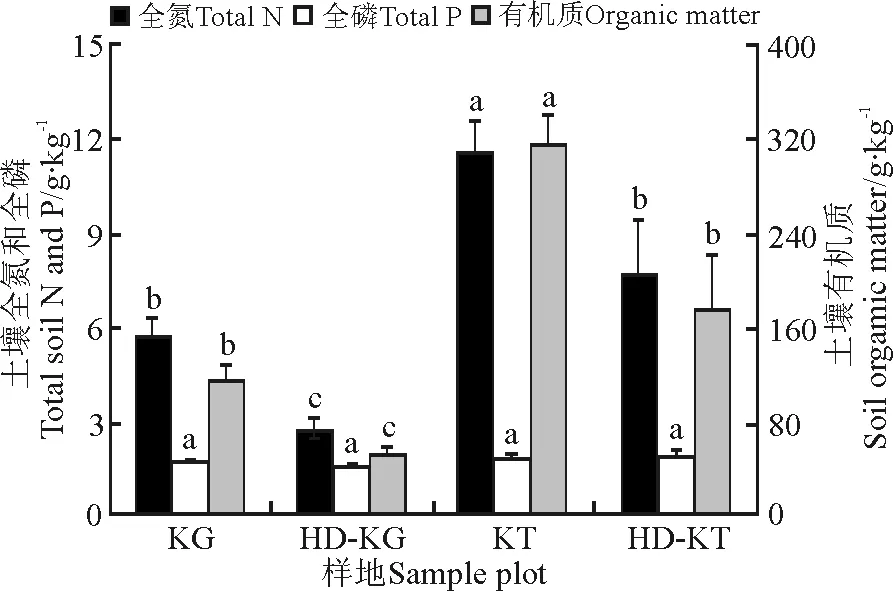
图2 不同样地土壤养分Fig.2 Soil nutrients of different sample plots
2.4环境因子逐步多重回归分析 不同因子的容差值不一致,土壤含水量、地上生物鲜质量、裸地面积、地上生物量、土壤全氮和土壤有机质的容差较小,表明这些因子具有较强的多重共线性;而土壤全磷、鼠洞密度、物种数和生草层厚度等容差值较大,表明这些因子受其它环境因子的影响较小(表2)。
2.5环境因子的相关性分析 优良牧草鲜质量与土壤含水量、生草层厚度、植被总盖度、植被高度、地上植物鲜质量、裸地面积、土壤全氮、土壤全磷和土壤有机质均呈极显著相关(P<0.01),R2分别为0.64、0.52、0.48、0.68、0.49、-0.46、0.51、0.48和0.64。
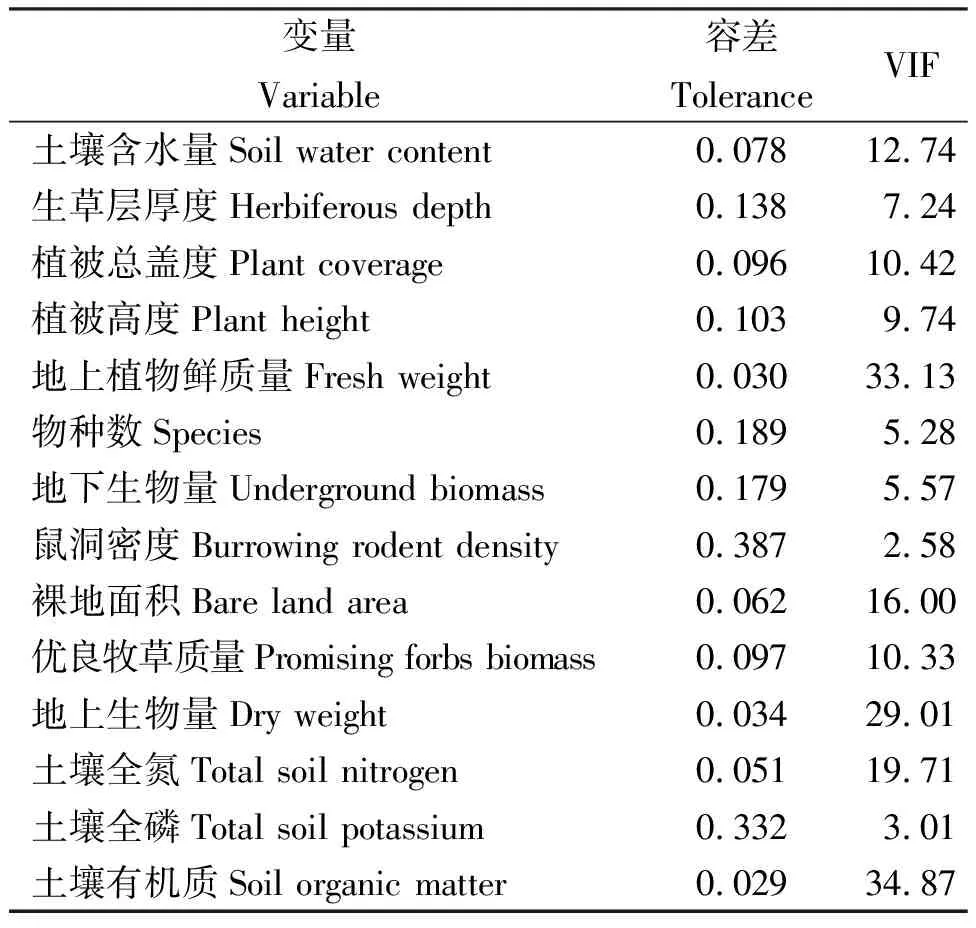
表2 环境因子容差分析Table 2 Analysis of environmental factors
2.6影响生物量的环境因子主成分分析 土壤养分因子对生物量的贡献率是不一致的,第1主成分与土壤含水量、土壤有机质、土壤全氮等呈显著正相关(P<0.05),表明土壤因子对生物量的影响程度较大。第2主成分与地上植物鲜质量、地上生物量、物种数和地下生物量呈显著相关,表明群落结构影响到生物量。第3、4主成分与裸地面积、生草层厚度、植被盖度和鼠洞密度呈显著相关,表明土壤基况、动物也一定程度地影响生物量的差异。但是,主成分1~5的累计贡献率也只是83.88%,表明除了土壤、群落结构等因子外,人为放牧、温度等因子对生物量的影响同样不容忽视(表3)。
3 讨论
3.1退化嵩草草甸优良牧草质量的变化与植物群落结构 衡量高寒草甸健康与否的重要指标之一是优良牧草质量。高寒嵩草草甸的退化首先表现为群落结构的演替变化[6],柳小妮等[7]在研究东祁连山退化草甸群落结构的变化中指出,随着退化程度的加剧,优良牧草逐渐演替成双子叶杂类草,优良牧草质量显著降低。赵玉红等[8]得出相似结论,退化草甸的优良牧草质量显著降低,杂类草逐渐成为退化草甸的优势物种。本研究表明,不论是小嵩草草甸还是藏嵩草草甸,退化后优良牧草质量都显著降低、杂类草质量增加(图1)、裸地面积增加、植被盖度降低(表1)。此外,由于双子叶杂类草的入侵,植被高度显著增加,而且变异程度也增高,这与于健龙等[5]的研究结果是一致的。

表3 土壤养分因子主成分分析矩阵Table 3 Principal component analysis matrix of soil nutrient factors
鼠害一直是危害高寒草甸的一个重要因子,长期在青藏高原高寒草甸生长的高原鼢鼠(Myospalaxrufescensbaileyi)和高原鼠兔通过直接采食优良牧草、掘土挖洞等活动极大地改变了草地植物量[9]。于健龙和石红霄[10]在研究高原鼠兔对高寒草甸植被的影响中指出,高原鼠兔严重破坏了高寒草甸植被群落结构,导致高寒草甸地上植物量显著降低。本研究表明,与健康小嵩草草甸、藏嵩草草甸相比,退化草甸的鼠洞密度均较高(表1),然而由于退化草甸植被群落结构的严重破坏,地上生物量显著降低(图1),缺少了高原属兔等所需的优良牧草,因此鼠洞密度变异程度增大(表1)。
3.2嵩草草甸的退化与土壤因子的关系 嵩草草甸的退化引起草地系统一系列的变化,包括植物物种结构[6]、土壤物理化学性质[11]、土壤酶[12]、土壤微生物[13]和土壤动物[14]等的变化。有研究表明,嵩草草甸群落地表植被与土壤因子间有强相关性[15]。王文颖等[16]研究指出,退化嵩草草甸的土壤有机碳、氮含量都显著下降,这在王长庭等[17]的研究中也得到证实。本研究中,退化嵩草草甸(小嵩草、藏嵩草)的土壤有机质、土壤全氮含量都显著下降,而土壤全磷则相对稳定(图2),土壤含水量下降(表1)。分析其原因,可能是土壤有机质主要来源于植物的枯枝落叶[18],而土壤全氮主要来源于土壤有机质[19],两者的矿化都受到土壤质地、水分以及温度的综合作用[20],因此草甸的退化引起土壤理化性质、土壤微环境的改变,从而加速土壤有机质和氮素的流失与土壤水分的蒸发,而土壤全磷则主要来自土壤母质[21],其变异系数较小,相对稳定。
近年来,关于退化嵩草草甸的治理模式研究已然成为热点[22],魏雪红等[23]通过建植栽培草地改良的方法成功改良了“黑土型”退化草甸。然而,关于如何治理退化嵩草草甸并未形成固定的模式,曹广民和龙瑞军[24]研究指出,三江源地区“黑土滩”退化草地并不能通过围封的方式自然恢复,必须通过建植栽培草地才能达到“黑土滩”的恢复。可见,探究退化草甸植被、土壤等众多因子的贡献程度将为开展退化草甸的人工恢复提供重要的科学依据。本研究发现,土壤含水量、土壤有机质和土壤全氮等有较强的多重共线性(表2),与优良牧草鲜质量存在显著正相关(表3)。主成分分析也表明,土壤因子是影响优良牧草鲜质量的主要因子之一(表3)。可见,土壤理化性质等土壤因子是决定优良牧草鲜质量的关键,除了土壤因子外,鼠洞密度、植被盖度等植被群落特征也对地上植物量有一定的贡献率(表3)。然而,本试验的主成分分析的所有因子的贡献率也只为83.88%,因此还有其它因子是本研究未涵盖的,如放牧[25]、光照[26]等都在一定程度上影响地上生物量的变化。
[1] 曹广民,林丽,张法伟,等.青藏高原高寒矮嵩草草甸稳定性的维持、丧失与恢复[J].草业科学,2010,27(8):34-38.
[2] 杨汝荣.我国西部草地退化原因及可持续发展分析[J].草业科学,2002,19(1):23-27.
[3] 马玉寿,郎百宁,李青云,等.江河源区高寒草甸退化草地恢复与重建技术研究[J].草业科学,2002,19(9):1-4.
[4] 李希来.高寒草甸草地与其退化产物“黑土滩”生物多样性和群落特征的初步研究[J].草业科学,1996,13(2):21-23.
[5] 于健龙,石红霄,李剑,等.不同退化程度高寒嵩草草甸基况的初步研究[J].草业科学,2010,27(9):115-118.
[6] 王炜,刘钟龄,郝敦元,等.内蒙古典型草原退化群落恢复演替的研究Ⅱ.恢复演替时间进程的分析[J].植物生态学报,1996,20(5):460-471.
[7] 柳小妮,孙九林,张德罡,等.东祁连山不同退化阶段高寒草甸群落结构与植物多样性特征研究[J].草业学报,2008,17(4):1-11.
[8] 赵玉红,魏学红,苗彦军,等.藏北高寒草甸不同退化阶段植物群落特征及其繁殖分配研究[J].草地学报,2012,20(2):221-228.
[9] 孙飞达,龙瑞军,郭正刚,等.鼠类活动对高寒草甸植物群落及土壤环境的影响[J].草业科学,2011,28(1):146-151.
[10] 于健龙,石红霄.高原鼠兔洞口密度对高寒嵩草草甸植被及土壤水分的影响[J].中国草地学报,2010(4):109-112.
[11] 赵云,陈伟,李春鸣,等.东祁连山不同退化程度高寒草甸土壤有机质含量及其与土壤主要养分的关系[J].草业科学,2009,26(5):20-25.
[12] 李以康,韩发,冉飞,等.三江源高寒草甸退化对土壤养分和土壤酶活性影响的研究[J].中国草地学报,2008,30(4):51-58.
[13] 于健龙,石红霄.高寒草甸不同退化程度土壤微生物数量变化及其影响因子[J].西北农业学报,2011,20(11):81-85.
[14] 吴鹏飞,杨大星.若尔盖高寒草甸退化对中小型土壤动物群落的影响[J].生态学报,2011,31(13):3745-3757.
[15] 王长庭,龙瑞军,王根绪,等.高寒草甸群落地表植被特征与土壤理化性质、土壤微生物之间的相关性研究[J].草业学报,2010,19(6):25-34.
[16] 王文颖,王启基,鲁子豫.高寒草甸土壤组分碳氮含量及草甸退化对组分碳氮的影响[J].中国科学D辑:地球科学,2009,39(5):647-654.
[17] 王长庭,龙瑞军,王启兰,等.三江源区高寒草甸不同演替阶段土壤有机碳和微生物量碳的变化[J].应用与环境生物学报,2008,14(2):225-230.
[18] 杨成德,龙瑞军,陈秀蓉,等.东祁连山不同高寒草甸类型土壤表层碳、氮、磷密度特征[J].中国草地学报,2008,30(1):1-5.
[19] 陈伟,杨国锋,赵云,等.金佛山地区不同生境下土壤有机质与全氮含量及其相关性[J].草业科学,2009,26(6):25-28.
[20] 王艳杰,邹国元,付桦.土壤氮素矿化研究进展[J].中国农学通报,2005,21(10):203-208.
[21] 刘兴诏,周国逸,张德强.南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征[J].植物生态学报,2010,34(1):64-71.
[22] 陈文业,郑华平,戚登臣.黄河首曲沙化草地恢复重建模式研究[J].草业科学,2008,25(6):14-18.
[23] 魏雪红,孙磊,武高林.青藏高原东缘“黑土型”退化草甸人工草地改良的土壤养分响应[J].水土保持学报,2010,24(5):153-156,168.
[24] 曹广民,龙瑞军.三江源“黑土滩”型退化草地自然恢复的瓶颈及解决途径[J].草地学报,2009,17(1):4-9.
[25] 王长庭,龙瑞军,王启兰,等.放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应[J].生态学报,2008,28(9):4144-4152.
[26] 冯超,古松,赵亮,等.青藏高原三江源区退化草地生态系统地表反照率特征[J].高原气象,2010,29(1):70-77.
