林分密度对水曲柳人工幼龄林植被碳储量的影响1)
2012-09-18刘婷岩张彦东彭红梅甘秋妹
刘婷岩 张彦东 彭红梅 甘秋妹
(东北林业大学,哈尔滨,150040)
森林是陆地生态系统的主体,具有较高的固碳和储碳能力,据估算森林生态系统每年固定的碳约占整个陆地生态系统的2/3[1],全球森林植被的碳储量约占全球植被碳储量的77%[2],森林生态系统较高的固碳和储碳能力对调节全球碳循环具有重要的作用。森林的固碳和储碳水平与森林的组成和结构密切相关[3],良好的森林结构是保证具有较高固碳和储碳能力的基础[4]。研究表明,随林分结构的改善,林分碳储量会相应的增加[5]。林分密度作为决定林分结构的重要因素,影响林分的碳储量[6-8]。目前,有关林分密度与植被碳储量的研究结果并不一致。有研究认为,随林分密度增加,植被碳储量增加[7,9-11];也有研究认为,随林分密度增加,植被碳储量降低[6];还有人认为,随林分密度增加,植被碳储量在各林分间无显著差异[12]。林分密度对森林植被碳储量的影响可能因树种、密度范围等不同而存在差异,深入的规律还需要进一步研究。
水曲柳(Fraxinus mandshurica)是我国东北东部地区珍贵硬阔叶树种之一,以往对水曲柳主要在适生立地[13-14],混交技术[15-16],密度与生长的关系[17]等方面开展了一些研究,而关于水曲柳人工林碳储量[18]方面的研究尚少。本研究以4种不同密度水曲柳人工林为对象,研究水曲柳人工林植被碳储量随林分密度的变化规律,为人工营造高固碳林分提供科学依据。
1 研究区概况
试验地设在东北林业大学帽儿山实验林场进德施业区,地理坐标为 127°30'~127°34'E,45°21'~45°25'N。该场位于长白山系张广才岭西坡,地貌类型为低山丘陵,平均海拔300 m。该区气候属大陆性季风气候,年平均气温2.8℃,年降水量723 mm,平均年蒸发量1094 mm,有效积温2526℃,无霜期120~140 d。地带性土壤为暗棕壤,非地带性土壤有草甸土、白浆土和沼泽土。地带性植被为阔叶红松(Pinus koraiensis)林,现存植被为原始植被经过反复破坏后形成的天然次生林和人工林。本区次生林类型多样,主要有蒙古栎(Quercus mongolica)林,水曲柳、胡桃楸(Juglans mandshurica)等组成的硬阔叶林,白桦(Betula platyphylla)、山杨(Populus davidiana)等组成的软阔叶林,以及槭树(Acer spp.)、椴树(Tilia spp.)、榆树(Ulmus spp.)等组成的杂木林等。本区人工林主要树种有落叶松(Larix gmelini)、樟子松(Pinus sylvestris)、红松、水曲柳和胡桃楸等。
2 研究方法
研究林分为13年生的水曲柳人工林,该林分为1998年春营造,共分4种密度,造林地为次生林的皆伐迹地。林分的初植密度分别为:2200株·hm-2(株行距为1.5 m×3 m)、2500 株·hm-2(株行距为2 m×2 m)、4400 株·hm-2(株行距为1.5 m×1.5 m)、10000株·hm-2(株行距为1 m×1 m),分别用Ⅰ、Ⅱ、Ⅲ、Ⅳ表示。不同密度的林分均位于东南坡的中下部,坡度<10°,坡面平整,立地条件相似。不同密度林分中,林下灌木和草本盖度存在差异,灌木盖度为2% ~5%,草本盖度为34% ~53%。林下灌木主要有:珍珠梅(Sorbaria sorbifolia)、卫矛(Evonymus sacrosancta)、悬钩子(Rubus crataegifolia)等。草本主要有:北重楼(Paris verticillata)、山尖子(Cacalia hasta)、铃兰(Convallaria majalis)、玉竹(Polygonatum odoratum)等。造林后林分一直未进行间伐,但因自然稀疏调查时林分密度已降低。调查时4种不同初植密度林分的现存密度如表1。
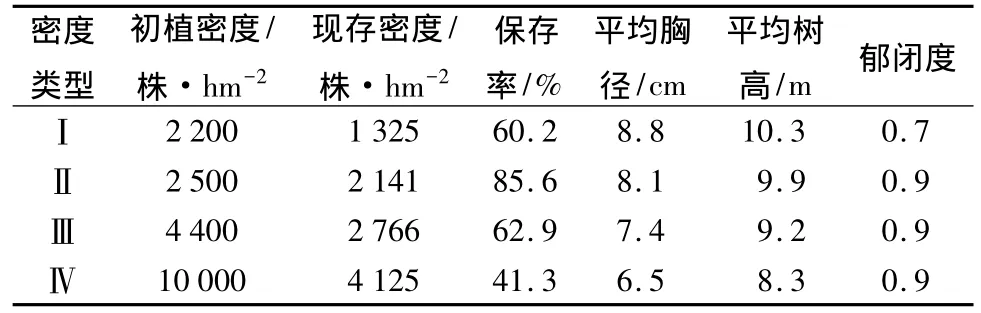
表1 不同初植密度水曲柳人工林的现存密度及基本状况
2.1 样地设置与样品采集
于2010年8月,在4种不同密度的水曲柳人工林内各设置3块20 m×20 m的样地进行调查。在已设置的样地内进行每木检尺,根据每木检尺的测定结果,在每块样地中各选择平均标准木1株,采用分层切割法,分别采集干、枝、叶样品。根系采用全根挖掘法,按直径大小分为大根(≥5 cm)、中根(2~5 cm)、小根(0.5~2 cm)3个等级采集样品,带回实验室。
在每块样地内按梅花形布设2 m×2 m和1 m×1 m的小样方各5个,分别进行灌木和草本生物量调查。分地上部分和地下部分,采用“样方收获法”全株收获样方内灌木和草本,测定鲜重。同时按地上和地下部分,分别采集灌木和草本样品,带回实验室。
2.2 样品处理与测定
将采集的乔木、灌木和草本样品在65℃烘箱中烘干至质量恒定后,用电子天平(±0.01 g)称质量,计算含水率。将采集烘干后的样品用高速粉碎机(天津市泰斯特仪器有限公司,FW177、FW100)磨碎,过筛后用元素分析仪(德国·Elementer,VARIO Macro)测定乔木不同器官、灌木和草本含碳量。水曲柳干、枝、叶和根的平均含碳量分别为:47.01%、48.28%、46.41%和47.70%,灌木地上部分和地下部分平均含碳量分别为44.46%和41.31%。草本地上部分和地下部分平均含碳量分别为41.19%和39.68%。
2.3 碳储量计算
乔木层碳储量计算:利用王传宽拟合的水曲柳生物量模型估算乔木层单株和林分生物量[19]。单株的碳储量即为单株各器官生物量与相应器官含碳量乘积的加和。林分碳储量即为所有单株碳储量加和,再换算成单位面积碳储量。
林下植被层碳储量计算:根据已测定的灌木、草本鲜质量和含水率,推算出各密度水曲柳人工林灌木和草本生物量。由灌木和草本地上和地下部分生物量和含碳量估算碳储量,再换算成单位面积碳储量。
2.4 数据分析
采用SPSS 18.0软件对数据进行统计分析,用单因素方差分析(ANOVA)检验不同密度水曲柳人工林碳储量之间的差异性,然后用最小显著差数法(LSD)检验差异显著性。
3 结果与分析
3.1 不同密度水曲柳人工林乔木层碳储量
林分密度对乔木层生物量和碳储量具有明显的影响,乔木层生物量和碳储量表现出随林分密度的增加而增大的变化趋势(表3)。在4种密度的林分中,乔木层生物量和碳储量由小到大的顺序为:Ⅰ<Ⅱ<Ⅲ<Ⅳ。经检验,密度Ⅰ的乔木层生物量和碳储量与密度Ⅲ、Ⅳ差异显著(P<0.05),其余密度间差异均不显著(P>0.05)。最高密度Ⅳ的生物量和碳储量比最低密度Ⅰ分别高出52.0%和52.1%。
乔木层各器官生物量和碳储量亦随林分密度发生明显变化(表3),干、枝、叶、根的生物量和碳储量均随密度的增加而增大。其各器官生物量和碳储量随密度变化的顺序与乔木层总生物量和碳储量变化相同。经检验,密度Ⅰ的干和根生物量和碳储量与密度Ⅲ、Ⅳ差异显著(P<0.05),其余密度间差异均不显著(P>0.05);密度Ⅰ叶生物量和碳储量与其它3种密度之间差异显著(P<0.05);枝生物量和碳储量在4种密度之间差异均不显著(P>0.05)。不论密度如何变化,乔木层各器官碳储量的分配均为干>根>枝>叶。乔木层各器官占总生物量和碳储量的比例随密度发生变化,干和叶所占比例随密度增加 而增大,而枝和根所占比例则随密度增加而降低。
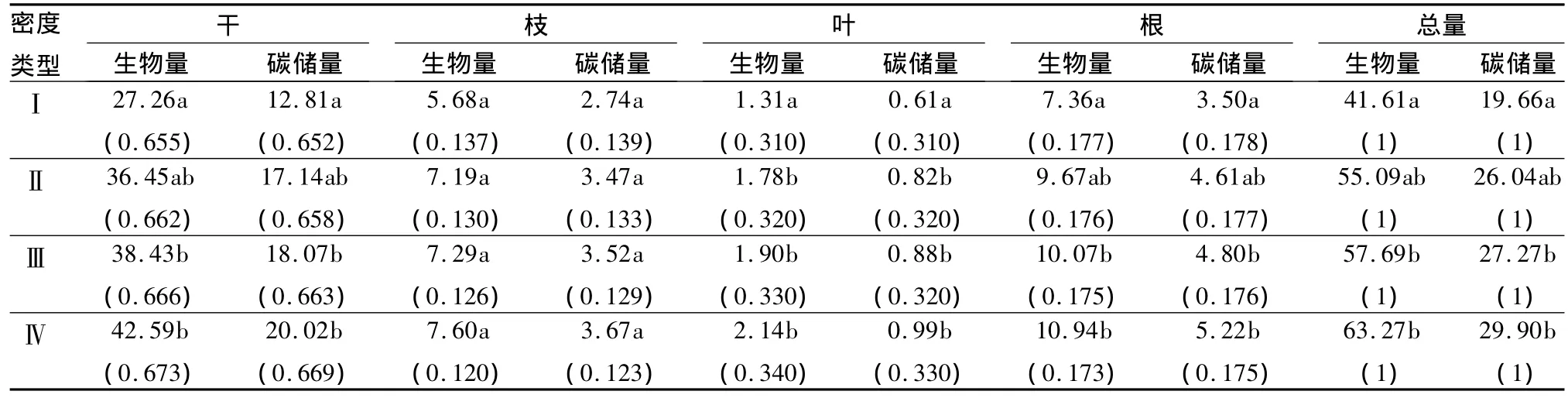
表3 不同密度水曲柳人工林乔木层碳储量 t·hm-2
林分密度对单株生物量和碳储量亦具有明显的影响,单株生物量和碳储量均表现出随林分密度的增加而减小的变化趋势(表4)。在4种密度的林分中,单株生物量和碳储量由大到小的顺序为:密度Ⅰ>Ⅱ>Ⅲ>Ⅳ。经检验,密度Ⅰ的单株生物量和碳储量与密度Ⅲ、Ⅳ差异显著(P<0.05),密度Ⅱ与Ⅳ差异显著(P<0.05),其余密度间差异均不显著(P>0.05)。最低密度Ⅰ的生物量和碳储量比最高密度Ⅳ分别高出105.9%和106.1%。
单株各器官生物量和碳储量随林分密度发生明显变化,干、枝、叶、根的生物量和碳储量均随密度的增加而减小(表4)。其各器官生物量和碳储量随密度变化的顺序与单株总生物量和碳储量变化相同。经检验,单株各器官的生物量和碳储量表现为,密度Ⅰ与密度Ⅲ、Ⅳ差异显著(P<0.05),密度Ⅱ与Ⅳ差异显著(P<0.05),其余密度间差异均不显著(P>0.05)。不论密度如何变化,单株各器官碳储量的分配均为干>根>枝>叶。各器官占单株总生物量和碳储量的比例随密度发生变化,干和叶所占比例随密度增加而升高,而枝和根所占比例则随密度增加而降低。
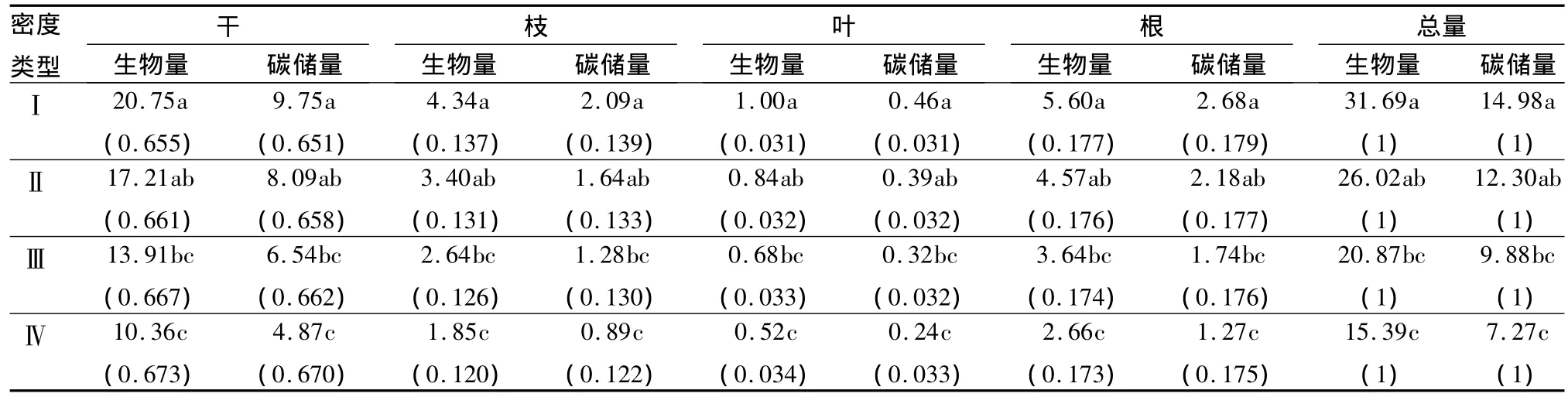
表4 不同密度水曲柳人工林乔木层单株碳储量 kg·株-1
3.2 不同密度水曲柳人工林林下植被层碳储量
林分密度对林下灌木层生物量和碳储量具有一定影响,灌木层生物量和碳储量均表现为在一定范围内随密度增加而减小,密度继续增加时基本不变的规律(表5)。经检验,密度Ⅰ的灌木层生物量和碳储量与其它密度差异显著(P<0.05),而其它3种密度间差异均不显著(P>0.05)。林分密度对草本层生物量和碳储量具有明显的影响,草木层生物量和碳储量均随林分密度增加而减小。经检验,密度Ⅰ的草木层生物量和碳储量与其它密度差异显著(P<0.05),密度Ⅳ与密度Ⅱ、Ⅲ差异显著(P<0.05),而密度Ⅱ和Ⅲ之间差异不显著(P>0.05)。
林分密度对林下植被层总生物量和碳储量亦具有明显的影响,林下植被层生物量和碳储量均随林分密度增加而减小(表5)。在4种密度的林分中,林下植被层生物量和碳储量由大到小的顺序为:密度Ⅰ>Ⅱ>Ⅲ>Ⅳ。经检验,密度Ⅰ的林下植被层生物量和碳储量与其它密度差异显著(P<0.05),密度Ⅳ与密度Ⅱ、Ⅲ差异显著(P<0.05),而密度Ⅱ和Ⅲ之间差异不显著(P>0.05)。最低密度Ⅰ的生物量和碳储量比最高密度Ⅳ分别高出84.2%和93.3%。
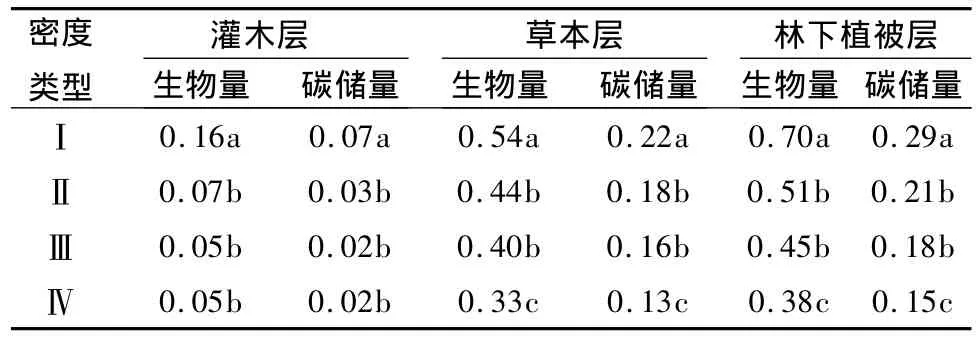
表5 不同密度水曲柳人工林林下植被层碳储量t·hm-2
3.3 不同密度水曲柳人工林植被碳储量
4种林分密度中,植被层碳储量由密度Ⅰ到密度Ⅳ分别为:19.95、26.25、27.45 和 30.05 t·hm-2,且随林分密度增加而增大(表6)。经检验,密度Ⅰ的植被层碳储量与密度Ⅲ、Ⅳ之间差异显著(P<0.05),其余密度间差异均不显著(P>0.05)。密度Ⅰ的植被层碳储量比密度Ⅲ、Ⅳ分别低37.6%、50.6%。不同密度林分中,乔木层碳储量均占植被总碳储量的绝大部分,比例达98.5% ~99.5%,且该比例随林分密度增加而增大。林下植被层碳储量占植被总碳储量的比例仅为0.5% ~1.5%,且该比例随林分密度的增加而减小。
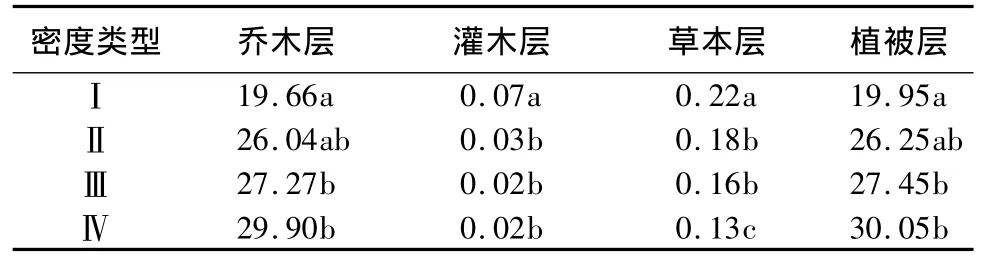
表6 不同密度水曲柳人工林植被碳储量 t·hm-2
4 讨论
不同密度的水曲柳人工林,随林分密度增加,乔木层碳储量增加,而林下植被层碳储量减小。在4种密度的林分中,乔木层碳储量为植被总碳储量的98.5% ~99.5%,均占有很高的比例。据热带、亚热带和温带等不同地区的研究,乔木层碳储量通常占植被层总碳储量的 95.3% ~99.5%[6-7,9],本研究结果与之吻合。这表明不同密度林分中,植被碳储量主要由乔木层碳储量决定。
水曲柳人工林植被碳储量随林分密度增加而增大,这与已有的多数研究结果一致[7,9-11],但也与一些研究结果不同[6,12]。潘辉等[6]对福建柏(Kokienia hodginsii)的研究认为,植被碳储量随林分密度增加而减小。而方晰等[12]对湿地松的(Pinus elliottii)研究认为,植被碳储量在不同密度间差异不显著。产生上述差异的原因可能与研究林分密度范围不同有关。研究表明,林分植被碳储量主要由乔木层碳储量决定,而乔木层碳储量与乔木层生物量成严格的比例关系[6-7,9],因此,可以认为密度对林分乔木层生物量与植被碳储量的影响规律是一致的。林分密度与产量的关系研究认为,林分存在一个合理密度范围,当林分处于合理密度范围内其生物量最高,低于合理密度下限和高于其上限林分生物量均降低[20],而且合理密度随林分年龄增加逐渐降低[21]。本研究中水曲柳林分年龄较小,现存密度仅为1325~4125株·hm-2。林分植被碳储量表现为随密度增加而增大,可能是4种林分密度均低于合理密度上限值。潘辉等[6]对福建柏的研究仅选用2种密度(1500和2725株·hm-2)林分,且林分年龄已达28年,其较低密度林分可能在合理密度范围内,而较高密度林分已经超出了合理密度上限,故表现为随密度增加植被碳储量降低。而方晰等[12]对16年生4种密度(2220、2745、3255 和3750 株·hm-2)湿地松林分的研究得出植被碳储量在各林分间无显著差异。可能是其4种林分密度均在合理密度范围内,故其碳储量无显著差异。林分密度对植被碳储量的影响因研究林分密度范围不同结果可能不同,选择由低到高多种密度的林分进行对比才会得到较完整的规律。但在多数研究中,林分密度常常处于有限的范围内,故得出的结论是多样的。
[1]樊后保,黄玉梓,袁颖红,等.森林生态系统碳循环对全球氮沉降的响应[J].生态学报,2007,27(7):2997-3009.
[2]Watson R T,Noble IR,Bolin B.Land use,land-use change,and forestry,Special Report of the Intergovernmental Panel on Climate Change.The United States of America:Cambridge University Press,2000.
[3]陈建成,程宝栋.森林与低碳经济刍议[C].南昌:第四届中国林业技术经济理论与实践论坛,2009:97-101.
[4]姚爱静.晋西黄土区林分结构特征研究[D].北京:北京林业大学,2005.
[5]巨文珍,王新杰,孙玉军.长白落叶松林龄序列上的生物量及碳储量分配规律[J].生态学报,2011,31(4):1139-1148.
[6]潘辉,赵凯,王玉芹,等.不同密度福建柏人工林碳储量研究[C].福州:经济发展方式转变与自主创新—第十二届中国科学技术协会年会,2010:1118-1122.
[7]王秀云,孙玉军,马炜.不同密度长白落叶松林生物量与碳储量分布特征[J].福建林学院学报,2011,31(3):221-226.
[8]杨晓菲,鲁绍伟,饶良懿,等.中国森林生态系统碳储量及其影响因素研究进展[J].西北林学院学报,2011,26(3):73-78.
[9]张国庆,黄从德,郭恒,等.不同密度马尾松人工林生态系统碳储量空间分布格局[J].浙江林业科技,2007,27(6):10-14.
[10]Fang S,Xue J,Tang L.Biomass production and carbon sequestration potential in poplar plantations with different management patterns[J].Journal of Environmental Management,2007,85(3):672-679.
[11]Fernández-Nú~nez E,Rigueiro-Rodríguez A,Mosquera-Losada MR.Carbon allocation dynamics one decade after afforestation with Pinus radiata D.Don and Betula alba L.under two stand densities in NW Spain[J].Ecological Engineering,2010,36(7):876-890.
[12]方晰,田大伦,项文化,等.不同密度湿地松人工林中碳的积累与分配[J].浙江林学院学报,2003,20(4):374-379.
[13]王庆成,张彦东,王政权.微立地土壤水分-物理性质差异及对水曲柳幼林生长的影响[J].应用生态学报,2001,12(3):335-338.
[14]范志强,沈海龙,王庆成,等.水曲柳幼林适生立地条件研究[J].林业科学,2002,38(2):38-43.
[15]张彦东,王庆成,谷艳华.水曲柳落叶松人工幼龄混交林生长与种间竞争关系[J].东北林业大学学报,1999,27(2):6-9.
[16]姜瑞凤,董哲,董恒.红松大青杨、红松水曲柳混交林试验效果分析[J].东北林业大学学报,2003.31(3):77-78.
[17]田宇明,王庆成,王鹏,等.水曲柳造林密度与林分生长的关系[J].林业科技,2009,34(5):17-20.
[18]梅莉,张卓文,谷加存,等.水曲柳和落叶松人工林乔木层碳、氮储量及分配[J].应用生态学报,2009,20(8):1791-1796.
[19]ChuankuanW.Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J].Forest Ecology and Management,2006,222(1-3):9-16.
[20]吴增志,杨瑞国,徐效智,等.杨树苗期合理密度规律及其在育林中的应用研究[J].河北林学院学报,1994,9(4):273-281.
[21]吴增志,徐效智,杨瑞国,等.杨树苗期合理密度与合理密度线研究[J].植物生态学报,1994,18(2):177-183.
