夹竹桃光合及荧光特性对海岸线距离的响应1)
2012-08-09吴统贵
董 毅 吴统贵
(中国林业科学研究院亚热带林业研究所,富阳,311400)
顾建明 虞木奎 成向荣 段 溪 王 臣
(上海市浦东新区林业站) (中国林业科学研究院亚热带林业研究所) (山东省乳山市林业局)
我国大陆海岸线北起辽宁,南至广西,长达18 340 km。沿海地区是我国经济最发达、社会发展最快的地区;同时,也因其特殊的地理位置以及气候条件,生态环境建设一直处在比较重要的位置。沿海防护林体系在抵御台风、风沙等自然灾害方面发挥着极其重要的作用,是我国沿海地区生态建设的重要内容。防护林树种除了受土壤盐碱影响以外,还遭受着诸如台风、泥石流、灾害性海浪等自然灾害的侵扰[1]。因此筛选出既耐盐碱又具有良好的抗风性树种显得尤为重要。
曾经有学者通过AHP的方法在上海地区2000多种树种中筛选出了适合在沿海地区推广种植的树种,包括木麻黄(Casuarina equisetifolia L.)、中山杉(Ascendens mucronatum)、海桐(Pittosporum tobira)、夹竹桃(Nerium indicum Mill.)等既耐盐碱又抗风的乔灌树种[2],并且相关的研究表明,筛选出的树种在盐碱与风环境中生长良好[3]。到目前为止有关沿海适生树种的抗性研究多集中在模拟控制试验,然而,由于树木在实地生长中受到诸多因素的影响,尤其沿海立地环境复杂多变[4-5],模拟控制试验往往不能说明树种在自然环境下的生理生态特性和环境适应性。因此,对具有特殊生境的沿海防护林进行实地研究具有重要的现实意义。
夹竹桃是龙胆目夹竹桃科的常绿直立灌木,具有耐污染性强、抗辐射强、环境修复能力强、经济价值高、观赏效果佳的特点,是我国东南沿海地区防护林广为种植的树种之一[6]。因此,本研究选取了距离海岸线不同距离(不同风速)的夹竹桃为研究对象,系统测定了东南西北4个方向上的叶片净光合速率、蒸腾速率、叶绿素荧光参数等,分析夹竹桃不同方向叶片生理生化特性在不同生长环境下的响应差异,以期为今后夹竹桃在沿海地区的推广种植和沿海防护林体系构建与改造提供一定的理论依据。
1 研究区概况
研究区位于上海市浦东新区,121°52'~121°57'E,30°58'~30°59'N,属亚热带湿润季风气候。年平均气温15~21℃,年平均降水量1 040 mm。由于是围垦形成的陆地且围垦年限不长,并没有形成典型的地带性植被,20世纪80年代中期沿着不同围垦年份的堤坝上构建水杉林带,至今已形成一定规模。海拔1~5 m。土壤多为淤泥质粉质黏土。在距离海岸线不同距离人工种植夹竹桃林带。
根据设在上海浦东新区滨海地区各个气象观测点所得的常年气象数据可知,林带最前沿的常年平均优势风向风速为8 m·s-1,在夹竹桃生长季盛行东南季风。
2 材料与方法
材料:2011 年 9 月,在距离海岸线 4、5、6、7、8 km处夹竹桃林带各选取3株长势良好的夹竹桃,分别按距离海岸线的近远标记为 D4、D5、D6、D7、D8等5个不同的点(见表1)。在每株东南西北4个方向上各选取3片代表叶进行光合参数和叶绿素荧光参数的测定。
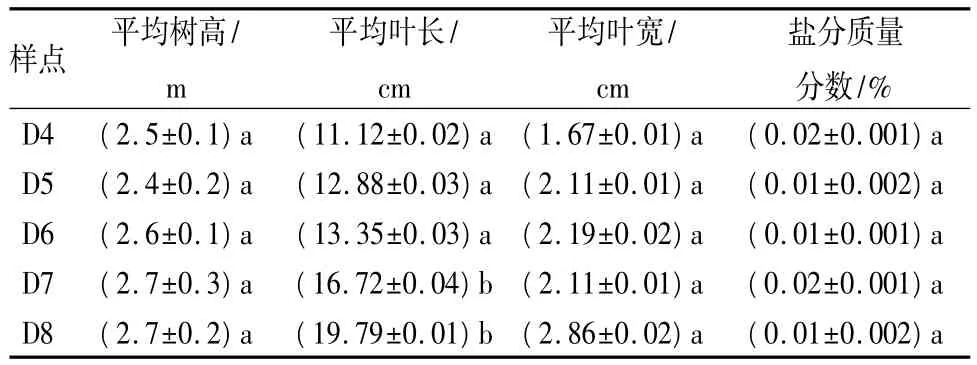
表1 距离海岸线不同距离的夹竹桃基本生长状况
光合参数的测定:每天的9:00—10:00,用 Li-6400便携式光合作用测定仪(Li-COR公司,美国)直接测定按不同距离、不同方向选取的叶片的净光合速率(Pn)和蒸腾速率(Tr),光合有效辐射控制在1 200μmol·m-2·s-1左右。所有测定重复3次,取平均值。
叶绿素荧光参数的测定:使用 PAM-2500(Walz,德国)便携式调制叶绿素荧光仪测定各项荧光参数,测定前先暗适应30 min,测定时调整叶片使其光量尽量一致,以减少误差。每天在9:00—10:00测定暗适应下最大荧光(Fm)、初始荧光(Fo)、PSⅡ最大光化学量子效率(Fv/Fm)、光合电子传递速率(ETR)、光化学猝灭系数(qP)、非光化学猝灭系数(qN)。所有测定重复3次,取平均值。
土壤盐分质量分数及电导率的测定:每处夹竹桃林带分别采用5点取样采集表层土,取回实验室后进行风干,研磨至粉状,用DDSJ-308A电导率仪进行测定,盐分质量分数在0.01%~0.02%,各点差异不大。
数据处理与分析:所有数据采用Excel2007处理完成,统计分析采用SPSS16.0统计分析软件中单因素方差分析(One-Way ANOVA)功能处理完成。
3 结果与分析
3.1 海岸线不同距离夹竹桃叶片光合、荧光特性变化规律
由表2可知,随着离海岸线距离的增加,夹竹桃叶片的净光合速率(Pn)和蒸腾速率(Tr)均呈现逐渐上升的趋势。D4作为滨海防护林体系前沿距离海岸线最近的一点,其夹竹桃叶片Pn((7.8±0.4)μmol·m-2·s-1)、Tr((0.6±0.1)mmol·m-2·s-1)显著低于其他各点(P<0.05)。
Fv/Fm是PSⅡ最大光化学量子产量,其大小反映了PSⅡ反应中心内初光能的转换效率(最大转换效率),该参数在植物受到胁迫时会明显下降,因此是反映光合器官受抑制程度的良好指标。如表2所示,随着距海岸线距离的增加,夹竹桃的最大光化学量子产量逐渐增加,且发现D4与D8之间存在显著差异(P<0.05)。
荧光猝灭分为光化学猝灭和非光化学猝灭两类。由表2可以看出光化学猝灭系数(qP)呈逐渐上升的趋势,而非光化学猝灭系数(qN)呈逐渐减小的趋势。但通过单因素方差分析发现各点之间差异并不显著(P>0.05)。
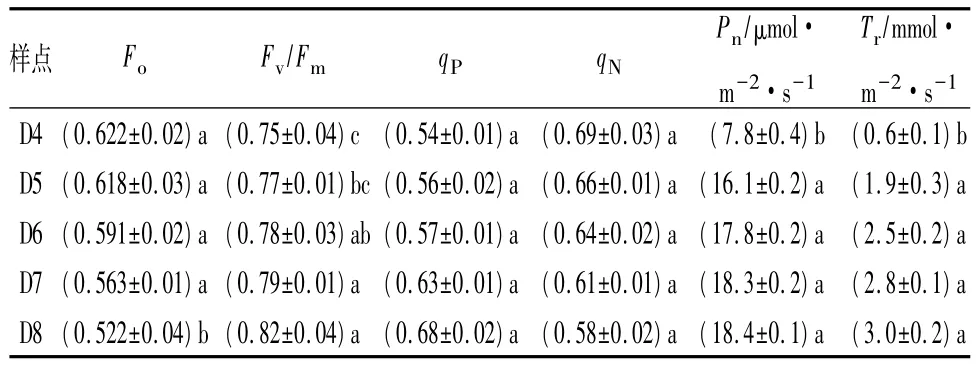
表2 距离海岸线不同距离的夹竹桃光合和荧光参数
3.2 不同方向夹竹桃叶片光和特性变化规律
由表3可以看出,各个方向之间的净光合速率并没有显著性差异(P>0.05)。东方向上夹竹桃叶片的 Pn((14.8±0.2)μmol·m-2·s-1)和 Tr((2.2±0.3)mmol·m-2·s-1)略微的低于其他3个方向的平均值。此外,南方向上的叶片的Pn与Tr((2.3±0.1)mmol·m-2·s-1)最高。同时可以看出,东方向叶片的qP显著小于其他方向(P<0.05)。而其他各荧光参数在各方向间差异不大。

表3 不同方向上夹竹桃叶片光合和荧光参数
4 讨论
4.1 距海岸线不同距离夹竹桃光合作用参数差异
本试验所取各条夹竹桃林带土壤盐分质量分数仅为0.01%~0.02%,远远低于盐胁迫(轻度盐化土壤的盐分质量分数为0.13%~0.22%)的水平[7],说明盐胁迫并不是对夹竹桃的生长造成影响的主要因子。
风是一个重要的环境因子,当风力达到一定程度时,会对植物的生长产生抑制作用。于云江等[8]通过对油蒿(Artemisia ordosica)进行不同程度风沙胁迫发现,风力越大,油蒿的净光合速率(Pn)降幅越大。这与本试验的结果相一致。但是,关于在风力胁迫下蒸腾速率的变化规律却存在一定的争议。蒸腾作用是随环境因子的变化能做出快速响应的生理生态过程[9]。于云江等[10]认为风能促进蒸腾,这是因为它们能将气孔外面的的水蒸气吹走,补充一些湿度相对较低的空气,使扩散层变薄或消失,外部扩散阻力减小,从而蒸腾速率加快。然而,Griddings[11]对于强风蒸腾做过一定研究,认为几乎所有强风后蒸腾都是发生于角质层,从而使气孔关闭、蒸腾速率下降。本试验中,D4处夹竹桃位于防护林带的前端,距离海岸线最近,所处环境的风速最大,此处夹竹桃叶片蒸腾速率值显著低于处于林带后的D5、D6、D7、D8处,说明 D4处夹竹桃在强风的影响下,气孔被迫关闭,从而避免风力太大而造成水分散失过多。而随着林带后风力的减弱,蒸腾速率又逐渐恢复,且呈上升的趋势。
4.2 距海岸线不同距离夹竹桃的荧光参数差异
在叶绿素荧光参数当中,叶绿素荧光的可变部分(Fv)与最大荧光值(Fm)的比值(Fv/Fm)可以反映开放的PSⅡ中心捕获激发能的效率,是植物胁迫的重要参数[12-15]。一般正常生长的植物其Fv/Fm值维持在0.8左右[16],只有受到抑制的情况下才会发生降低现象[17-18]。徐娟娟等[19]通过研究荒漠藻类植物在风沙胁迫下的光合活性发现,风速为7 m·s-1条件下荒漠植物的Fv/Fm明显低于风速为3 m·s-1。本实验中,D4处夹竹桃的Fv/Fm平均值明显低于其他4处(P<0.05),说明叶片的PSⅡ反应中心在风胁迫的影响下受到破坏或者失活。同理,Fo值的增加也印证了PSⅡ反应中心受到破坏或失活。
光化学猝灭系数反映了PSⅡ天线色素吸收的光能用于光化学传递的份额,非光化学猝灭系数反映了PSⅡ天线色素吸收的光能不可以用作光合电子传递而以热的形式耗散掉的光能部分。光化学猝灭系数(qP)的降低反映了PSⅡ反应中心失活使得天线色素原初捕获光能力减弱,植株吸收的光能减少。但由于电子传输途径受到损伤,光合电子传递能力减弱,使得用于光化学反应的光能部分显著减少,从而形成过剩光能。当大量的过剩光能不能及时用于光化学反应时,就能形成光抑制现象,植物为了免受PSⅡ受到破坏,往往通过叶黄素循环、荧光发射、光呼吸等热耗散途径来消耗过剩光能[20]。本实验中,受风力影响较大的D4处夹竹桃叶光化学猝灭系数显著低于D8。另外,反映光能用于热耗散程度的非光学猝灭系数(qN)虽然没有呈显著性差异,但是从表2中可以看出,用于热耗散的光能呈下降的趋势,说明随着风力胁迫的减轻,植株将所获得的光能更多地用于光化学过程[21-22]。
4.3 不同方向上夹竹桃叶片的光合作用和叶绿素荧光特性的差异
光合器官是植物的最敏感部位,叶绿素α荧光与光合作用的各种反应密切相关,包含了很多光合作用的信息,逆境因子对植物光合作用的影响也可通过它反映出来[23-24]。本实验中,东向上的叶片作为生长季时主要的迎风方向,长期处于风力胁迫的影响下,与其他方向上的叶片相比,其受胁迫程度本应该更大。但是通过差异显著性检验(LSD)表明,除光化学猝灭系数(qP)呈显著小于其他方向外,其他参数所得出的值之间差异均不显著。这说明东方向上的叶片的电子传递活性由于长期受到5级(8 m/s)风的胁迫,已经形成适应性的降低。
夹竹桃叶片的PSⅡ反应中心开放程度比较小,电子传递活性也发生了一定程度的减弱。但是其生长并没有因为在单方向受到胁迫而呈现出显著的生长差异。这可能是由于:在风力胁迫下夹竹桃形成了一定的生态适应机制[25-26],或由于夹竹桃特殊的树形所致。因为夹竹桃并没有明显的主干,而成伞状多分枝生长,湍流动能经过夹竹桃时可能已被重新分配,从而造成风力对各个方向的生长影响差异不大[27]。这些还有待做进一步的探讨。
5 结论
本研究结论表明,风力的胁迫是造成夹竹桃的生长受到抑制的主要因素之一,且随着风力的加强,其受到抑制的程度越高。而风力并未对不同方向上叶片生长特性带来一定的差异。这些可能是由于夹竹桃特殊的树形结构将来自优势方向的风动能平均分配的结果导致的。总体而言,夹竹桃已经适宜了在风环境下生长,适合继续推广栽植。
[1]许景伟,李传荣.沙质海岸防护林体系构建技术研究[M].北京:中国林业出版社,2009:1-15.
[2]杨学军,郑天祎,储亦婷,等.上海地区沿海防护林树种的筛选应用[J].防护林科技,2003(4):5-7.
[3]陈胜.沿海防护林优良适生树种造林效果研究[J].南京林业大学学报:自然科学版,2005,29(4):91-95.
[4]成向荣,虞木奎,张建锋,等.沿海防护林工程营建技术研究综述[J].世界林业研究,2009,22(1):63-67.
[5]Ulanova N G.The effects of windthrow on forests at different spatial scales:A review[J].Forest Ecology Management,2000,135:155-167.
[6]杨淑贞.红花夹竹桃防护效能与育苗造林技术[J].防护林科技,2000(4):65-66.
[7]张建锋.盐碱地生态修复原理与技术[M].北京:中国林业出版社,2007:15-60.
[8]于云江,史培军,鲁春霞,等.不同风沙条件对几种植物生态生理特征的影响[J].植物生态学报,2003,27(1):53-58.
[9]王铸豪.植物与环境[M].北京:科学出版社,1986:60-64.
[10]于云江,史培军,贺丽萍,等.风沙流对植物生长影响的研究[J].地球科学进展,2002,17(2):262-266.
[11]Griddings L A.Transpiration of silpbium Laciniatam[J].Plant world,1914,35:937-942.
[12]李敦海,宋立荣,刘永定.念珠藻葛仙米叶绿素荧光与水分胁迫的关系[J].植物生理学通讯,2000,36(3):205-208.
[13]王良桂,张春霞,彭方仁,等.干旱胁迫对几种楸树苗木叶片荧光特性的影响[J].南京林业大学学报:自然科学版,2008,32(6):119-122.
[14]吕洪飞,皮二旭,王岚岚,等.遮荫处理的白英光合作用和叶绿素荧光特性研究[J].浙江师范大学学报:自然科学版,2009,32(1):1-6.
[15]胡学华,蒲光兰,肖千文,等.水分胁迫下李树叶绿素荧光动力学特性研究[J].中国生态农业学报,2007,15(1):75-77.
[16]梁芳,郑成淑,孙宪芝,等.低温弱光胁迫及恢复对切花菊光合作用和叶绿素荧光参数的影响[J].应用生态学报,2010,21(1):29-35.
[17]刑庆振,郁松林,牛雅萍,等.盐胁迫对葡萄幼苗光合及叶绿素荧光特性的影响[J].干旱地区农业研究,2011,29(3):96-100.
[18]王文杰,李文馨,祖元刚,等.紫茎泽兰和叶片色素及叶绿素荧光相关参数对不同温度处理的响应差异[J].生态学报,2009,29(10):5424-5432.
[19]徐娟娟,张德禄,吴国樵,等.风力胁迫对具鞘微鞘藻结皮光和特性的影响[J].水生生物学报,2010,34(3):575-580.
[20]Donald R O.When there is too much light[J].Plant Physiology,2001,125:29-32.
[21]柯世省,金则新.水分胁迫和温度对夏腊梅叶片气体交换和叶绿素荧光特性的影响[J].应用生态学报,2008,19(1):43-49.
[22]Coutts M P,Grace J.Wind and Trees[M].Cambridge:Cambridge University Press,1995.
[23]Krause G H.Cholophyll fluorescence and photosynthesis[J].Annual Reivew of Plant Physiology and Plant Moleculular Biology,2006,140:220-235.
[24]郑秋玲,谭伟,马宁,等.钙对高温下巨峰葡萄叶片光合作用和叶绿素荧光的影响[J].中国农业科学,2010,43(9):1963-1968.
[25]温莉娜,江波,杜天真,等.SO2对夹竹桃生理生化的影响[J].浙江林业科技,2008,28(3):59-62.
[26]Dunham A R,Cameron D A.Crown,stem and wood properties of wind-damaged and undamaged Sitka spruce[J].Forest Ecology and Management,2000,135:73-81.
[27]Zhou X H,Brandle J R,Takle E S,et al.Estimation of the three-dimensional aerodynamic structure of a green ash shelterbelt[J].Agricultural and Forest Meteorology,2002,111(2):93-108.
