温度和盐度对褶皱臂尾轮虫复合类群休眠卵体积的影响
2012-06-06王珊赵文焉鸿启
王珊,赵文,焉鸿启、2
(1.大连海洋大学水产与生命学院辽宁省水生生物学重点实验室,辽宁大连116023;2.沈阳市水产科学研究所,辽宁沈阳110034)
长期以来,褶皱臂尾轮虫Brachionus plicatilis被认为是广泛分布于内陆和沿海咸水水体的单一物种[1],包括S-型和L-型两种类型。随着科学家对生物复合类群研究的深入和分子生物学手段的介入,他们发现褶皱臂尾轮虫应该被视为一个包含隐匿同胞物种的轮虫复合类群 (B.plicatilis complex),其中至少包括3个物种[2-4]。这一复合类群可以根据其主要的进化关系及体型分为三大类群——大型 (L)、中型 (SM)和小型 (SS)。2001年,Ciros-Pérez等[5]分别详细描述了这三大类群中3种轮虫 B.plicatilis Müller、B.ibericus n.sp.和B.rotundiformis Tschugunoff的基本形态特征、生态特点和交配行为,并且使用分子生物学方法确定了其种类。这一复合类群会随着季节的改变发生明显的周期性变形。目前,相关的研究基本都集中在食物密度对休眠卵大小的影响上[6-10],而关于轮虫休眠卵的体积随周围环境的变化而发生改变的研究未见报道。为了更好地了解轮虫同胞物种间的生态差异和季节变异的机理,本研究中,作者在新分类标准的基础上,研究了不同温度和盐度对褶皱臂尾轮虫复合类群中3种同胞物种轮虫Brachionus plicatilis Müller、B.plicatilis Yamamoto 和 B.rotundiformis Tschugunoff休眠卵体积变化的影响。
1 材料与方法
1.1 材料
试验用褶皱臂尾轮虫复合类群采自大连市自然博物馆附近的一个沿海半咸水池塘——明珠花园水池 (体积约8886 m3,盐度为5)。
1.2 方法
1.2.1 轮虫的采集和预培养 试验前,先将轮虫在光照培养箱 (Thermo-3744,±0.1℃)内驯化培养7 d,培养温度为20℃,盐度为5,光周期为12L∶12D,光照强度为2500 lx。培养用水取自黑石礁海区,经煮沸消毒后,用蒸馏水稀释到需要的盐度,用VIR-211型盐度计在20℃下测定海水盐度。每天投喂蛋白核小球藻Chlorella pyrenoidosa,每周更换一次培养液。从驯化后的轮虫中挑选单只轮虫单独培养,使其经孤雌生殖形成种群。
1.2.2 种类鉴定 从培养出的每个轮虫种群中随机挑选出3只带卵的孤雌生殖个体,用体积分数为5%的甲醛固定后,在Olympus-CX21显微镜下进行镜检。将具有典型种类特征的3种轮虫挑选出来,分别培养,种类鉴定参照前人的标准[5,11-13]。各种轮虫经过孤雌生殖培育形成群体后,将单只轮虫饥饿后进行DNA提取、PCR扩增、COⅠ测序,参照董云伟等[14]的方法,将测序结果与GenBank中已发表的相关序列进行比较,分别应用DNAstar 5.0和ClustalX 1.81进行比对,并确定其种类。扩增引物参照Folmer等[15]设计的引物,每个类群的检测设3个重复。
1.2.3 温度和盐度对3种轮虫休眠卵体积的影响试验 随机从 3种轮虫 B.plicatilis Müller、B.plicatilis Yamamoto和B.rotundiformis Tschugunoff(分别简称为Bm、By和Br)中挑选活力强的怀卵非混交雌体放入烧杯中培养,培养液体积为50 mL,每个烧杯中放25只轮虫。温度分别设为15、20、25℃,每个温度下分别设置3个盐度 (5、15、30)。每个试验设2个重复。
试验在光照培养箱 (Thermo-3744,±0.1℃)中进行,光周期为12L∶12D,光照强度为2500 lx,投喂蛋白核小球藻,饵料密度为2×106个/mL。培养用水取自黑石礁海区,经煮沸消毒后,用蒸馏水稀释到需要的盐度。每6 h搅动一次培养液,使饵料悬浮有利于轮虫摄食,每2 d更换一次培养液。试验时间为15 d,15 d后收集休眠卵。将收集的休眠卵用体积分数为5%的福尔马林固定,并在Olympus-CH21型显微镜 (15×40)下测量轮虫的卵长 (L,μm)和卵宽 (W,μm),每组测量20个休眠卵。休眠卵体积 (V,μm3)按照以下公式计算[7]:
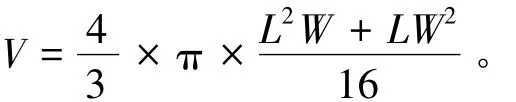
1.3 数据处理
使用SPSS 11.5软件对数据进行单因素方差分析和三因素方差分析,用Duncan法进行多重比较。
2 结果
2.1 种类鉴定
对3种轮虫形态学的测量结果表明:Bm的平均被甲长和被甲宽分别为270.90、219.80 μm,By的平均被甲长和被甲宽分别为269.20、205.95 μm,相似的同胞物种Bm的体型比By稍大;Br的体型最小,平均被甲长和被甲宽分别为153.08、124.81 μm。对3种轮虫的被甲长和被甲宽进行方差分析后可知,3种轮虫之间存在极显著差异 (n=25,P< 0.01)。
将通过形态学鉴定为 B.plicatilis Müller、B.plicatilis Yamamoto和B.rotundiformis Tschugunoff的线粒体COⅠ序列在GeneBank中进行比对,结果发现:Bm的COⅠ区序列与AY785178(Brachionus plicatilis isolate tianjin4)和AP009408(Brachionus plicatilis mitochondrial)DNA的COⅠ区序列覆盖率和相似度都为100%,说明该种轮虫确为B.plicatilis Müller;By的COⅠ区序列和AP009408(Brachionus plicatilis mitochondrial)的COⅠ区序列覆盖率为97%,相似度为98%,说明该种轮虫为B.plicatilis Müller的同胞物种;Br的COⅠ区序列和AY785225的COⅠ区序列覆盖率和相似度均为100%,说明该种轮虫确为B.rotundiformis Tschugunoff。
2.2 休眠卵体积
将3种轮虫的休眠卵体积进行三因素方差分析(tree-way ANOVA)和多重比较,结果表明,温度、盐度、种类及温度与盐度的交互作用对轮虫休眠卵的体积影响均极显著 (P<0.001)。
从表1可见:在各种温度和盐度条件下,Br休眠卵的体积均显著小于另外两种轮虫同胞物种Bm和By(P<0.05),而这两种体型相近的轮虫Bm和By休眠卵的体积只在高温低盐 (25℃,5)、高温中盐 (25℃,15)、中温中盐 (20℃,15)和中温高盐 (20℃,30)的条件下差异显著(P<0.05)。Br休眠卵的平均体积为 3.68×105μm3,而Bm和By休眠卵的平均体积分别为10.88×105μm3和10.69×105μm3。3 种轮虫中,休眠卵平均体积最小的是Br,为2.99×105μm3,出现在低温高盐 (15℃,30)条件下;休眠卵平均体积最大的是By,为13.23×105μm3,出现在低温低盐(15℃,5)条件下。
当温度为15~25℃时,Br和Bm的休眠卵体积在各盐度下均呈现先升高后下降的趋势,而By的休眠卵体积则呈现先下降后升高的趋势。当盐度为5~30时,3种轮虫休眠卵体积在各温度下呈现的变化规律不太一致,总体上呈下降的趋势。
各温度条件下,Bm和By休眠卵的体积差异显著 (P<0.05)。在中温条件下,Br的休眠卵体积比其它温度条件变化显著 (P<0.05),而低温和高温条件下的Br休眠卵体积之间变化不显著 (P>0.05)。
在低盐条件下,Bm和By的休眠卵体积比其它盐度条件变化显著 (P<0.05);在高盐条件下,Br的休眠卵体积比其它盐度条件变化显著 (P<0.05)。
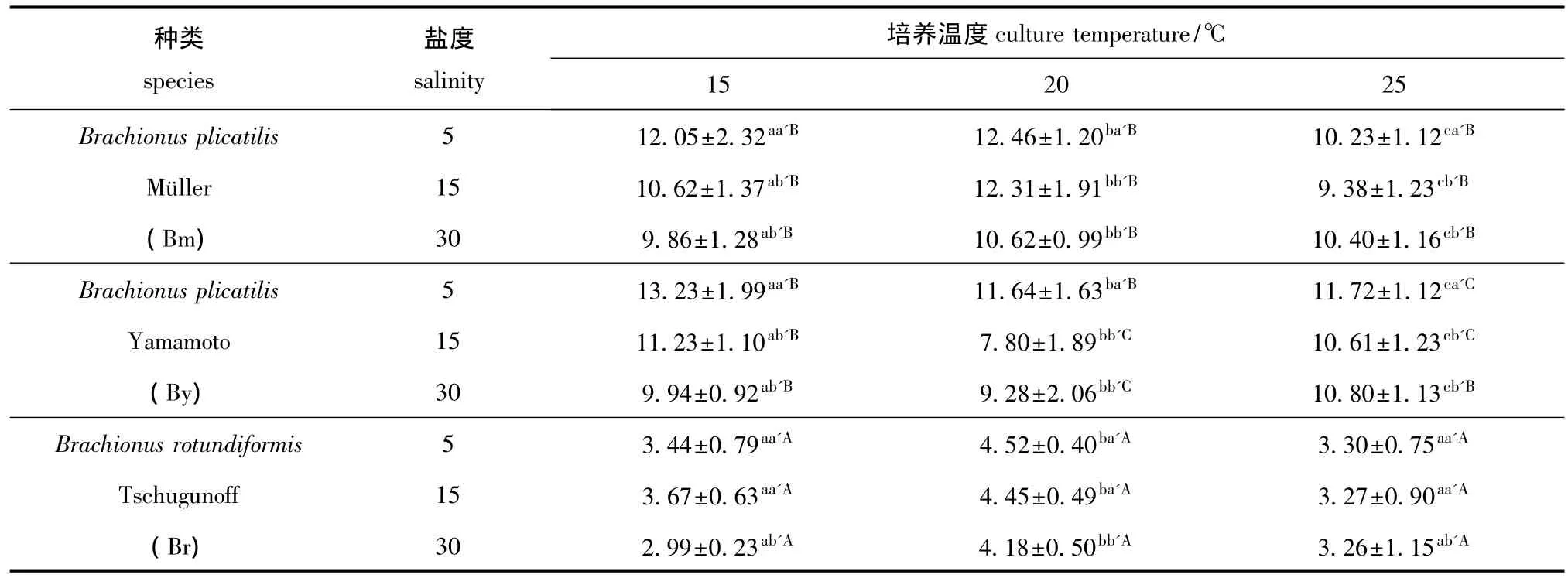
表1 不同温度和盐度下3种轮虫休眠卵的体积Tab.1 The resting egg volume of three rotifer species at different temperatures and salinity 105μm3
3 讨论
本研究中鉴定出的Bm和By在基因和表型上都具有很大的相似性,在低温 (15℃)条件下这两种轮虫休眠卵的体积差异并不显著,可能与它们在低温条件下基础代谢降低有关。但在温度较高的条件下,两种轮虫休眠卵的体积却表现出了不同的变化规律,说明微小的基因差异就有可能导致其在不同环境条件下产生不同的变化。温度、盐度及其交互作用对这两种相似的同胞物种休眠卵的体积影响显著 (P<0.05),其中盐度和温度与盐度的交互作用对其影响极显著 (P<0.01),说明对于这两种L-型轮虫来说,休眠卵体积对盐度的改变较温度更为敏感,这与它们生活的半咸水池塘的盐度常年保持在5有关。
自然条件下,同一水域分布的轮虫休眠卵的体积会有很大差异[16],且沉积在自然水域底泥中的褶皱臂尾轮虫休眠卵体积大小会随着底泥垂直分层的不同而不同[17]。这种现象可能与环境因子对轮虫休眠卵体积的影响有关。为了探究这一现象,人们研究了不同温度和盐度条件下轮虫休眠卵的变化,结果表明,在极端的温度和盐度条件下,褶皱臂尾轮虫的休眠卵体积会较大[18];Pavón-Meza等[19]在对轮虫Brachionus havanaensis的研究中也证明休眠卵体积与轮虫体型呈线形相关。本研究中,Bm和By的休眠卵体积在各个温度和盐度条件下都明显大于Br休眠卵的体积,这与此3种轮虫的个体大小是紧密相关的:Br本身属于S-型轮虫,而Bm和By则属于L-型轮虫。本研究中,最大的平均休眠卵体积出现在低温低盐 (15℃,5)条件下,而在此条件下Bm和By休眠卵的体积偏大(表1)。由于Br对低温 (15℃)不适应性,这种轮虫在低温条件下没有表现出这一特点,但在相对适应的温度和盐度条件下——中温低盐 (20℃,5),该轮虫休眠卵体积最大。这说明尽管在极端温度和盐度条件下轮虫休眠卵的体积会有所变化,但较大的休眠卵只会出现在该种轮虫较为适应的环境中。
[1]Walker K F.A synopsis of ecological information on the saline lake rotifer Brachionus plicatilis Müller 1786[J].Hydrobiologia,1981,81-82:159-167.
[2]Gómez Á,Temprano M,Serra M.Ecological genetics of a cyclical parthenogen in temporary habitats[J].Journal of Evolutionary Biology,1995,8(5):601-622.
[3]Gómez Á,Serra M.Behavioral reproductive isolation among sympatric strains of Brachionus plicatilis Müller 1786 insights into the status of this taxonomic species[J].Hydrobiologia,1995,313 -314:111-119.
[4]Gómez Á,Snell T W.Sibling species and cryptic speciation in the Brachionus plicatilis species complex(Rotifera)[J].Journal of Evolutionary Biology,1996,9(6):953-964.
[5]Ciros-Pérez J,Gómez Á,Serra M.On the taxonomy of three sympatric sibling species of the Brachionus plicatilis(Rotifera)complex from Spain,with the description of B.ibericusn sp.[J].Journal of Plankton Research,2001,23(12):1311-1328.
[6]Kirk K L.Egg size,offspring quality and food level in planktonic rotifers[J].Freshwater Biology,1997,37:515-521.
[7]Sarma S S S,Rao T R.Effect of food level on body size and egg size in a growing population of the rotifer Brachionus patulus Müller[J].Archivfür Hydrobiologie,1987,111:245-253.
[8]Walz N,Rothbucher F.Effect of food concentration on body size,egg size and population dynamics of Brachionus angularis(Rotatoria)[J].Verhandlungen des Internationalen Verein Limnologie,1991,24:2750-2753.
[9]胡好远,席贻龙,耿红.食物浓度对角突臂尾轮虫种群增长、个体大小和卵大小的影响[J].应用生态学报,2002,13(7):875-878.
[10]耿红,席贻龙,胡好远.食物种类和浓度对红臂尾轮虫种群增长、个体大小及卵大小的影响[J].应用生态学报,2003,14(5):573-576.
[11]Sudzuki M.Intraspecific variability of Brachionus plicatilis[J].Hydrobiologia,1987,147:45-47.
[12]Segers H.Nomenclatural consequences of some recent studies on-Brachionus plicatilis(Rotifera,Brachionidae)[J].Hydrobiologia,1995,313-314:121-122.
[13]殷旭旺,赵文.沿海混盐水体褶皱臂尾轮虫的复合类群及其种群动态[J].大连水产学院学报,2005,20(1):1-10.
[14]董云伟,牛翠娟,鲍蕾,等.单只轮虫 DNA提取及其细胞色素C氧化酶:I亚基部分序列测定[J].动物学研究,2002,23(1):81-83.
[15]Folmer M,Black W,Hoeh R,et al.DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrate[J].Molecular Marine Biology and Biotechnology,1994,9:294-299.
[16]Gilbert J J.Dormancy in rotifers[J].Transactions of the American Microscopical Society,1974,93:490-513.
[17]Snell T W,Burke B E,Messur S D.Size and distribution of resting eggs in a natural population of the rotifer Brachionus plicatilis[J].Gulf Research Reports,1983,7(3):285-287.
[18]Serrano L,Serra M,Miracle M R.Size variation in Brachionus plicatilis resting eggs[J].Hydrobiologia,1989:186-187:381-386.
[19]Pavón-Meza E L,Sarma S S S,Nandini S.Combined effects of temperature,food(Chlorella vulgaris)concentration and predation(Asplanchna girodi)on the morphology of Brachionus havanaensis(Rotifera)[J].Hydrobiologia,2007,593(1):95-101.
